Studying mammals: A winning design
Use 'Print preview' to check the number of pages and printer settings.
Print functionality varies between browsers.
Printable page generated Tuesday, 9 June 2026, 10:22 AM
Studying mammals: A winning design
Introduction
Mammals come in a bewildering variety of shapes and sizes and yet all of the 4700 or so species have some characteristics in common. Indeed, it's the existence of these common features that justifies the inclusion of all such diverse types within the single taxonomic group (or class) called the Mammalia.
This is the first in a series of units about studying mammals. To get the most from these units, you will need access to a copy of The Life of Mammals (2002) by David Attenborough, BBC Books (ISBN 0563534230), and The Life of Mammals (2002) on DVD, which contains the associated series of ten BBC TV programmes. You should begin each course by watching the relevant TV programme on the DVD and reading the corresponding chapter in The Life of Mammals. You will be asked to rewatch specific sequences from the programme as you work through the course.
This OpenLearn course provides a sample of level 1 study in Environment & Development
Learning outcomes
After studying this course, you should be able to:
explain the distinctive biological features of monotremes
distinguish contrasting modes of reproduction in monotremes, marsupials and placental mammals
describe the cellular basis of lactation and explain the benefits of an early diet of milk
explain the significance of mammalian metabolic rate
explain how and why the thermogenic response differs among species.
1 Overview
As you work through this course you will come across boxes, like the one below, which give you advice about the study skills that you will be developing as you progress through the course. To avoid breaking up the flow of the text, they will usually appear at the start or end of the sections.
As well as the course text, you will be using The Life of Mammals book (LoM) and related The Life of Mammals DVDs, as described in the introduction to this course. Before you go any further, watch 'A Winning Design' on the DVD and read LoM Chapter 1.
To gain maximum benefit from the question that follows the next paragraph, make an attempt at answering the question before you reveal the answer. You will probably find it helpful to write down an answer to the question, in note form at least, and then compare it with the one given.
Mammals come in a bewildering variety of shapes and sizes and yet all of the 4700 or so species have some characteristics in common. Indeed, it's the existence of these common features that justifies the inclusion of all such diverse types within the single taxonomic group (or class) called the Mammalia.
Question 1
On the basis of your study of LoM Chapter 1 and the associated TV programme, list up to three biological features that define mammals.
Answer
Mammals are distinguished by:
the production of milk;
the possession of hair (or similar structures, e.g. bristles);
and (along with birds) the ability to regulate their body temperature, usually being warm-blooded.
For much of this course, you will be concerned with exploring these diagnostic features of mammals in more detail. Sections 2 and 3 explore topics that emerge from the reading and viewing you've done already on the less 'conventional' mammals - the monotremes and the marsupials. Many of the facts and ideas in these early sections should therefore be familiar, though you'll also encounter new technical terms, especially in Section 3. One new issue that will come up a good deal is how we should best use the terms 'primitive' and 'successful'.
The title of this course emphasises the notion of mammalian design. This concept is a useful 'hook' on which to hang some introductory ideas; for example, how the structure of a particular part of an animal - a bone or a limb, perhaps - relates to its function, but biologists worry that using the term might imply that something complex has to be designed. (If you work through the series of units, you'll appreciate why any such implication is so strongly resisted.) Another idea emphasised is the success of mammals. You'll recall images from the TV programme of mammals flourishing in seemingly hostile environments. The fact that mammals are very widely distributed - such that David Attenborough (from here on referred to as DA) has to travel from the Arctic to the scrubland of Australia to sample mammalian diversity - is an expression of their success as an animal group. Another useful measure might be numbers of species - though the 4700 or so species of mammal is dwarfed in comparison with the known number of insect species in the world, which some estimate to be as many as 10 million. Other expressions of the success of mammals that are worth considering include the degree of physiological sophistication of their systems (which can be difficult to assess) and the number of individuals of a particular species or broader group that exist in total.
Mammals have also been around for a long time; LoM [p. 14] mentions the shrew-like 200-million-year-old fossil named Megazostrodon. Rather than shuffling along, with splayed-out limbs in the manner of many reptiles, this animal had limbs that were more erect and aligned under the body. Fossil evidence shows that the skulls of very early mammals have a distinctive lower jaw structure and sites on the skull for the attachment of chewing muscles. We can be confident that between 225 and 195 million years ago, mammal-like reptiles evolved into true mammals, though for the next 100 million years or so these unobtrusive animals, none larger than a pet cat, continued (in DA's words) to 'scuttle around the feet of the dinosaurs'. Their diversity did not appear until more than 100 million years later, during a period of geological time that witnessed the demise of the dinosaurs and their close relatives.
2 How should we think of monotremes?
This section contains the first of the activities, Activity 1. If possible, you should do each activity as you come to it; the text that follows it assumes you have done so. However, if this is not possible, then try to do the activity at the first opportunity, rather than leaving all the activities to do together at the end of the course. The activities usually need you to write something down and you will probably find it best to use a (hard-backed) notebook for your notes to keep them safe and together. Activity 1 asks you to watch a sequence on the DVD and make some notes from it on a particular subject. Pause the DVD if you need to, while you write things down. Don't spend too much time on this and don't worry if you find it difficult - you will get a lot more practice later on if you work through all the units in the series. Again, keep the answer concealed while you are doing the activity.
Recall the 'stars' of the early part of LoM and the TV programme - the echidna and the duck-billed platypus. These modern mammals are so distinct that they are assigned to a discrete subclass, the Prototheria, which includes the order Monotremata, separate from the more familiar and well-studied mammals that occupy most of our attention in this series of units. In LoM [p. 20] these two animals are termed 'part-reptile, part-mammal', a phrase that we'll look at more critically in a moment.
Activity 1
Watch 'A Winning Design' on the DVD from 06.32-17.40 and briefly write down the mammalian features evident in these monotremes. Is their egg-laying habit similar in all ways to that displayed by reptiles, such as snakes?
Answer
Both echidnas and the platypus have fur - though in the former, some of the hairs are thickened in the form of spines.
Both animals produce milk - they have mammary glands, though well defined nipples are not evident.
Both are 'warm-blooded' - but you'll see in Section 5 why we should be wary of using this everyday term too freely.
The egg-tooth in the hatchling [p. 17] is reminiscent of reptiles, but immature young hatch from their soft-shelled eggs after about 10 days; snakes typically hatch in a much more mature state. Thereafter, a young monotreme, though no longer carried, remains dependent on the mother's milk - for more than six months in the case of echidnas. You'll have watched that wonderful sequence of DA peering electronically into a platypus' breeding burrow, with an immature newborn obviously dependent upon prolonged maternal support.
Years ago, biologists often thought of the term 'egg-laying mammal' as synonymous with 'reptile-like mammal' or 'primitive mammal'. Now, with our greater understanding of monotreme biology, these emotive terms are disapproved of, since these animals have so many authentic mammalian features. For example, if echidnas didn't lay eggs, you might be forgiven for thinking of this animal as (in DA's description [p. 16]) 'little more remarkable than a rather large and slightly chilly hedgehog'; the hedgehog is a 'true' (or placental) mammal, as you'll see if you study the next course. I've mentioned that the period of development within the egg is relatively brief, but many aspects of reproduction and maternal care in the monotremes are distinctly mammalian.
Neither monotreme species is genuinely primitive or unsuccessful. They are specialist feeders; the platypus feeding on invertebrates (e.g. 'freshwater shrimp, an insect larva or a small mollusc' [p. 18]) living in the bottom of streams, while echidnas are terrestrial carnivores. The word 'primitive' implies a similarity with ancestral types, but in terms of lifestyle and anatomy, there's not a lot to link monotremes with the ancestral mammals typified by Megazostrodon.
There are further reasons not to think of monotremes as ineffective species that haven't quite 'made it' in an evolutionary sense. As part of their specialist forms of feeding, monotremes have highly developed sense organs - remember the 'remote-sensing' device of the platypus [p. 18]. If we think of numbers as a measure of an animal's success, monotremes are certainly successful. Echidnas - at least the short-nosed species - are described as 'quite common across their geographical range'. The platypus inhabits a particular type of environment (i.e. it occupies an environmental niche) that is threatened by human habitation, so numbers fell soon after the beginnings of European settlement in Australia in the 18th century. Recent conservation measures have meant that the species is no longer under severe threat.
And how long have monotremes existed? The platypus fossil that DA shows us in the TV programme (see 18.51), and mentions in LoM [p. 20], is about 25 million years old, but the oldest monotreme fossil (a jaw bone) is over 100 million years old. Their evolution could therefore be described as conservative, with little evidence of major changes over time. However, biologists remain ignorant about what the ancestors of monotremes looked like and what historic relationship they had with the evolutionary lines that gave rise to marsupial and placental mammals.
In the section you have just read, there were a number of references to pages in LoM given in square brackets [ ]. Indeed, unless stated otherwise, all the page references you encounter will be to LoM. Sometimes you may feel that you remember enough and do not need to go back and look again. If a particular species is mentioned, you may want to find more information about it. Use the index of mammals in LoM [pp. 316-320] to look it up. For example, echidnas were mentioned earlier in this section. Page 317 of the index gives you its scientific name, Tachyglossus aculeatus, together with references to text on p. 14 and illustrations (page numbers given in bold type) on pp. 15 and 17.
3 Reproduction in marsupials
The study of mammals requires you to deal with measurements, which we call numerical 'data', and you will get practice with compiling and analysing data if you work through all the units in this series. We assume only that you can add, subtract, multiply and divide. In this section, we ask you to use units - grams and kilograms, abbreviated to g and kg, respectively - and to calculate a percentage, for which you will probably need to use a calculator.
In contrast to monotremes, no marsupial lays a shelled egg. You'll know from LoM and the DVD that the embryo develops for a short period inside the uterus (or womb) before transferring to (in most species) a pouch; hence marsupials are sometimes termed 'pouched mammals'. The newborn are tiny and very unlike the adult - so much so, that the description of 'little worms' for the numbat [p. 21] is understandable, though some newborns are considerably more developed, as evident from p. 28. An adult female koala might tip the scales at about 8 kilograms (kg), but the newborn koala weighs just about half a gram, i.e. 0.5 g. Just how tiny this newborn is in relation to the mother becomes clearer if we work out what percentage its weight is of the mother's weight. But to compare 'like with like' we first need to express each measurement in the same units, in this case grams (g). 8.0 kg is equivalent to (8.0 × 1000) grams = 8000 g, so 0.5 g as a percentage of 8000 g is (0.5/8000) × 100%. This calculation comes to a little over 0.006% - compared to the mother, the newborn koala is very small indeed!
Question 2
Suppose a human baby weighs 3.4 kg at birth. If the mother weighs 70 kg, express the newborn's weight in relative terms, i.e. as a percentage of the mother's weight, to the nearest whole number.
Answer
Here the units are identical, so you just need to divide the baby's weight by that of the mother, 3.4/70, and multiply by 100%, which gives a value close to 5%. This is a great deal higher than the value for the koala, reflecting the greater relative maturity of the human newborn.
Activity 2
Watch 'A Winning Design' on the DVD, from 20.53-24.04, which focuses on birth in grey kangaroos. Jot down in your notebook the most striking points about the birth.
Answer
I was struck by the 'incredible journey' from birth pore to pouch and by the youngster's ability to orientate itself and to move with the aid of well-developed forelimbs. (Though perilous, the journey is relatively brisk - two minutes in all.) I was reminded of the importance of milk to the newborn and noted the fact that the chemical composition of the milk changes during the newborn's development.
After reading LoM, you'll be alert to the danger of thinking of the marsupial method of reproduction as 'primitive' or inferior to that of placental mammals. What we see is a successful reproductive strategy (or rather a range of strategies, because the details vary between different marsupial species) that is very different from our own. What is biologically so interesting is that such a large fraction of the early development of the young occurs after birth, in the pouch.
Question 3
LoM pp. 29-31 suggests that kangaroos 'have brought the marsupial method of reproduction to its most efficient level'. Can you recall what remarkable events offer evidence for such a claim?
Answer
LoM describes a very productive and seemingly appropriate method of producing young. Recall what was said on pp. 31-32 about the three young of the kangaroo, each at very different stages of development. In addition to the dormant or developing embryo in the uterus, the mother can support a youngster suckling in the pouch, plus yet another offspring that is largely independent but, as LoM points out [p. 32], returns to feed from time to time, for milk of a particular composition. (The timings of reproduction described at this point in LoM refer to the continuous breeding of the red kangaroo; the details of reproduction vary considerably amongst the 14 species within the subfamily that comprise the kangaroos, wallaroos and wallabies.)
To describe fully these complex events requires some new vocabulary. In many kangaroos, females mate very soon after giving birth. In the event of conception, the tiny ball of dividing cells, called a blastocyst, stops developing after a few days and the process of attachment to the inner lining of the uterus is prevented. In most forms of mammalian reproduction, a blastocyst would undergo such implantation without significant delay - indeed, in humans it's seen as marking the beginnings of true pregnancy. But in kangaroos the blastocyst remains 'frozen in time' in what is technically termed embryonic diapause. Some time just before the youngster in the pouch is ready to leave, the blastocyst implants and development proceeds to the point of birth. At about that point, the mother actively encourages the older offspring to spend less time in the pouch and prepares the pouch for the new arrival. Soon after the birth, mating is likely to lead to a further conception, and so on.
It's impossible to know for sure why embryonic diapause evolved, but the link with drought mentioned by DA [p. 32] is plausible. (If you work through the units in this series, you'll encounter many speculative explanations of unconventional mammalian strategies, trying to explain what selective advantage they might confer; if this mode of thinking is unfamiliar to you, then don't worry - it will be covered later.) During a severe drought, a suckling young in the pouch may be expelled and the heavy energetic demands of producing milk temporarily suspended. As DA points out [p. 32], this reduced energy expenditure will 'make the minimum demand on the meagre pasture around her and she has a fertile dormant egg within her womb ready to start its development just as soon as conditions improve'.
4 Milk production (lactation)
In this section, you are presented with a fairly complex diagram, Figure 1. The first thing to do when you come to any diagram is to read the caption (i.e. the title), which explains what it shows. Then look at the diagram itself, taking particular note of the scale, if there is one. You may find that the accompanying text 'talks you through' the diagram step by step, as it does here. Visualising three-dimensional objects from a two-dimensional diagram is a difficult skill and you should not expect to master it straight away, so don't be concerned if you don't immediately understand all aspects of Figure 1.
I'll now say more about one of the defining features of mammals - milk production. This feature isn't talked about in any detail in LoM but it's such a remarkable and unique mammalian process that it warrants attention here. The production of milk (technically called lactation) makes sense only if we look inside mammals to find out how this life-sustaining substance is produced.
Milk is a very rich form of food. You've probably already heard about some of the major constituents of milk - proteins, fats and carbohydrates. These large molecules have to be built up (synthesised) from the simpler chemicals that the mother obtains from her diet or from her body reserves. By looking at the structure of a typical mammary gland we'll see how this biological 'production line' is put together.
The term 'gland' is used for specialised structures that produce (or more technically secrete) one or more chemical products, and many glands have the type of structure that Figure 1 shows. (Glands are usually made up of different types of cells - a group of cells that have similar structure and function is often called a tissue.)
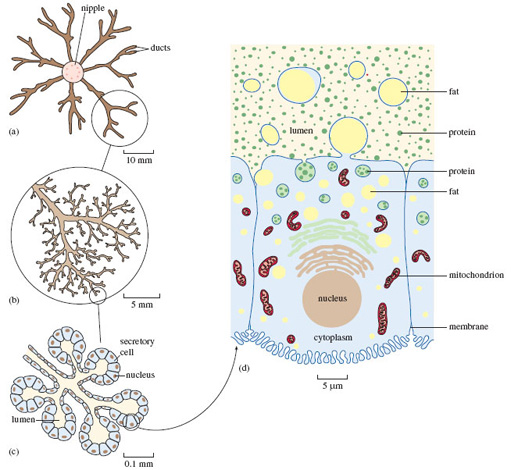
Figure 1a shows that each mammary gland consists of a central teat or nipple, into which feed a number of channels (or ducts) that convey and temporarily store the milk, following its production by the great mass of cells that make up the bulk of the gland. We get a better sense of the fine-detailed structure of the mammary gland by magnifying just one part of what's shown in (a). You'll appreciate that all the ducts shown have much the same structure, but focusing on just one makes things clearer. Figure 1b shows one representative part, around the blind ends of the finely branched ducts. At this magnification, the individual secretory cells aren't visible; but you can make them out in Figure 1c, a higher-powered view of just one small part of (b). In (c) you can see five such small groups, shown in a schematic way in cross-section - in other words, it's a 'tidied up' view of an imaginary slice though the gland. If you imagine these structures in three dimensions, (c) would resemble a 'bunch of grapes', the stalks representing the ducts and each 'grape' a group of secretory cells surrounding a central space (or lumen), which would contain milky fluid.
The structure of each individual cell in (c) is much the same and resembles that of many other cells in the mammalian body. Each cell is roughly rectangular in cross-section, with a thin outer membrane. Towards the base of the cell is a small rounded nucleus, which contains most of the genetic material (the DNA), which has a key role in directing the workings of the cell.
Figure 1d is a hugely magnified view of just one of the secretory cells shown in (c) - a single sample of the many millions of cells that comprise the mammary gland. Magnification on this scale requires the use of an electron microscope, as opposed to the less powerful optical light microscope used for (a)-(c) that you might be more familiar with. You'll recognise the cell's roughly rectangular shape and the rounded nucleus; you can also see the cell membrane.
The identity and function of all the many different cellular components in (d) needn't concern us in detail. There are many flattened, highly folded membranes, resembling stacked piles of plates. A few of these are shown in (d), just above the nucleus. (You will notice that there are two types of internal membrane stacks, denoted here in different colours.) The sausage-shaped structures are called mitochondria (singular: mitochondrion) - often termed the powerhouses of the cell, because they deliver the energy that fuels the complex synthetic processes that I'll talk more of in a moment. Towards the upper part of the cell, closest to the lumen, there are a number of fluid-filled droplets (called vesicles), containing what look like small granules. In reality, they are mostly different types of protein - just one of a range of large molecules (i.e. macromolecules) that make up the chemical constituents of all living material. In fact, I've drawn attention to proteins in Figure 1d by showing them as granules; though even at this level of magnification, proteins would not be discernible, since they are soluble in the fluid that contains them. Milk contains many proteins and most of them are assembled into these complex structures from much simpler chemical building blocks (called amino acids) within the secretory cells like the one shown in (d). The proteins in milk are vital to the growth and wellbeing of the suckling and some (collectively called antibodies) help the youngster withstand infection.
Fat is another key constituent of milk. You'll know that the fat content of milk can vary over time [p. 32], and the variation in milk composition between species is even greater. For example, rhinoceros milk contains hardly any fat, while seal milk is almost 50% fat. In those species where milk production has been most thoroughly investigated, secretory cells in the mammary glands take up fats from the bloodstream. (They also synthesise fats from other nutrient molecules carried to them in the blood, such as sugars.) The basic constituents of milk fats are then assembled together, most of them in parallel stacks of folded membranes of the type shown in Figure 1d. Fat accumulates within the droplets evident in (d), in the process of migrating to the upper part of the cell. The fat droplet, itself wrapped in a membrane, merges with the cell membrane and is 'budded-off' - fat droplet plus its enveloping membrane - into the lumen. From (d) you'll appreciate how each such structure resembles a decorative ring, with a fragment (of cell) often protruding from the central sphere - hence the name 'signet'.
Figure 1d conveys a sense of great cellular activity - all these synthetic processes require building blocks and the input of energy. These demands explain why lactation is a very considerable 'investment' by the mother in the wellbeing of her offspring. For animals on an unreliable or low-energy diet - the koala is a good example of the latter - synthesis of the constituents of milk can be a precarious operation. Given that the energy (fat) content of koalas' milk is comparatively low, it's not surprising that the period of lactation in koalas is unusually long, even by marsupial standards. At the age of five months, young koalas start to ingest eucalyptus leaves that have been already partly digested by the mother.
Lactation in marsupials has a particular importance; for example, the newborn red kangaroo (like the koala newborn mentioned in Section 3) weighs less than a gram - or, in the more familiar language of the TV commentary, 'less than a lump of sugar'. On complete emergence from the pouch, some eight months later, it weighs about four to five kilograms. It may then often double in weight before becoming fully independent of the mother's milk (i.e. becoming weaned), which happens between four and eight months after leaving the pouch. The composition and flow of milk are tightly controlled; the synthesis of particular nutrients can become switched on as particular genes are activated. (Such genes comprise particular sections of DNA within the nucleus of secretory cells, so minute that not even the magnification of Figure 1d can reveal them.) Towards the end of its time in the pouch, when the rate of the youngster's growth is very high and energy demands greatest, the fat content of the milk in many species reaches a maximum.
At a stage in development where hair growth is important, the youngster's production of the protein keratin has to be stepped up - keratin is the key ingredient of hair and hair-like structures. At such a time, the kangaroo's milk contains large amounts of the particular types of amino acid that are essential for the synthesis of keratin.
Question 4
Kangaroos and wallabies generally have four teats within the pouch, though there is only one newborn at a time. Why does the possession of more than one teat make sense?
Answer
Because such species have a unique and sophisticated system that allows the production of more than one type of milk, so satisfying the differing demands of youngsters at different stages of development [p. 32]. The mechanisms in the mother's body are not well understood but having separate 'plumbing' to each teat means each can act partially independently.
There's no doubt that the evolution of lactation was crucial to the success of mammals. Of course, milk production is just one component of a 'package' of parental care measures shown by many mammals, so it's perhaps risky to single out a particular feature, but it's one that raises particular interest.
Question 5
Can you think of some advantages of the lactation habit in mammals?
Answer
Lactation provides nutritional care - the sucklings don't have to find their own food. In that sense, it helps separate the infant from the vagaries of the environment. Given that feeding is relatively effortless, the youngster's energy can be largely channelled into growth. It also seems that suckling is an excellent way of transferring immunity from mother to offspring. You may know that the early milk of many mammals, humans included, confers a special advantage - it's especially rich in antibodies. The same is true in marsupials such as kangaroos. Clearly, the young are also protected in a variety of ways; often the protection is physical - remember the suckling marsupials from the TV programme.
5 Metabolism and body temperature
5.1 Warm-blooded vs cold-blooded
This section is concerned with another key mammalian characteristic. LoM highlights some of the essential terms and ideas - such as the notion of warm-bloodedness - but for more thorough study, some technical background is needed.
Activity 3
Watch the 'A Winning Design' on the DVD from 00.00-06.25, which emphasises the diversity of mammalian habitats. Using that information, plus your recollection of the LoM chapter, suggest some benefits of 'warm-bloodedness' to mammals and some of the implications of this strategy.
Discussion
Mammals are no longer at the mercy of the environment; by contrast, the body temperature of most amphibians, for example, is closely linked to outside temperatures. Thus the arctic fox remains warm and active in very cold conditions. But being warm-blooded has a high energetic cost. (In mammals, as much as 80-90% of the energy obtained from their food is needed to maintain body temperature.)
LoM p. 12 points out that non-mammals generate body heat, giving examples of a python and some species of fish, such as tuna. A reptile basking in the sun can become as warm to the touch as a mammal. If you've encountered a mammal during hibernation - perhaps a cautious investigation of a hedgehog - you'll probably have found it surprisingly cold to the touch. So in describing what's special about the body temperature of mammals, cold-blooded and warm-blooded are terms best avoided.
5.2 Metabolism
Scientific data are often presented in the form of a table, with the data arranged in columns (running vertically) and rows (running horizontally). You will find Table 1 below. As with diagrams, a table has a title and you should read that carefully. Look at the headings of the rows and columns. The units in which the values are measured are usually given in the column (or row) headings, rather than being written beside each value.
Metabolism is the technical name given to the sum total of all the chemical transformations inside cells. Many such changes involve building up complex chemicals - for example, the proteins and fats I've already mentioned - from simpler building blocks. In animals, the energy that such 'building-up' processes require has to come from the process of breaking down foodstuffs - the progressive breaking down in our bodies of the complex macromolecules that comprise our diet. (The final stages of breakdown and release of useful energy take place in the mitochondria, like those you saw in Figure 1d.) None of these energy-yielding or energy-consuming chemical transformations is 100% efficient and inevitably heat is released as a by-product. So an animal with a high level of metabolism, i.e. a high metabolic rate, produces a greater amount of heat than one operating at a lower level.
The chemical transformations we've talked of depend ultimately on the animal's intake of oxygen. In very simplified terms, oxygen is used in the mitochondria to complete the final stages of the breakdown (or more technically, the oxidation) of small energy-rich molecules - products of the chemical fragmentation of macromolecules mentioned earlier. In this process, usable energy (and some heat) are released.
Measuring how much oxygen is consumed by an animal over a period of time, such as a minute or an hour, is a pretty good measure therefore of the intensity of metabolism (i.e. the metabolic rate). But if we simply measure oxygen consumption in two animals that are very different in size - say a mouse and an elephant - we'd find that overall the elephant consumed much more oxygen over a minute than did a mouse, simply because a massive elephant has so many more cells in which metabolism is whirring away. If our interest is in metabolic rate, we'd have to take into account differences in the size by calculating the volume of oxygen consumed for a particular amount (or mass) of animal. Volumes of a gas such as oxygen, would be measured in millilitres (ml) or, more usually, the numerically equivalent unit, cubic centimetres (cm3), and the mass of the animal in grams (g), or perhaps in thousandths of a gram, i.e. milligrams (mg). In practical terms, oxygen consumption by a mouse of known mass would be measured over a period of time, say 15 minutes, and the values recalculated as so many 'cubic centimetres of oxygen per gram of mouse per hour'. (A more scientifically correct way of expressing the same thing is cm3 O2 g−1 h−1, and I'll have more to say about using this type of scientific notation in S182_3 Studying mammals: chisellers) In principle, the calculation for the elephant would be done in the same way, though one suspects with a few practical problems along the way.
| Metabolic rate/cm3 O2 g−1 h−1 | ||
|---|---|---|
| Species | at rest | at peak activity |
| salmon | 0.08 | 0.60 |
| monitor lizard | 0.08 | 0.38 |
| turtle | 0.03 | 0.64 |
| humming-bird | 2.80 | 42.00 |
| mouse | 2.50 | 20.00 |
| dog | 0.33 | 4.02 |
| human | 0.23 | 3.20 |
Footnotes
You might notice from these few data an implication that small mammals, e.g. the mouse, have a higher metabolic rate than larger mammals, such as the human. This trend is borne out by many other measurements, but I'll ignore it for the moment and return to it in S182_2 Studying mammals: the insect hunters.)Question 6
Look at Table 1, above, which shows the metabolic rates of a number of animals. What is the most striking difference between the values for mammals and those for fish, reptiles and a bird? (Concentrate on the 'at rest' values.)
Answer
The metabolic rates for mammals (and the humming-bird) are much higher than those for the fish and reptiles. This is true of both the 'at rest' levels and, as you'll see if you now look at the far right column, also the 'peak activity' values; the mouse, dog and human (and the humming-bird) values are higher than the remainder, often considerably so.
5.3 Heat production
There are two graphs in this section, Figure 2 and Figure 3. Graphs are often used to display data in a way that makes trends easier to see than in a table. Simple graphs have two axes, one running horizontally across the page, and the other vertically up the page. They are labelled in the same way as the column headings of a table (see Section 5.2) and the data points are then plotted. The text here talks you through that process. You will also notice the use of a type of scientific 'shorthand' in this section, where letters are used to represent words. The italic letter T is often used to represent 'temperature'. Here both the temperature inside an animal's body and the temperature of the surroundings are measured, so we need to identify which temperature we are referring to. This is done by adding an appropriate letter, usually subscripted and not italicised, immediately after the T. So here Tb stands for the body temperature of the animal and Ta for the surrounding, or ambient, temperature. When you see the shorthand version, try to translate it in your head into its real meaning, so I would read Tb as 'body temperature' rather than as 'tee-little-b'.
The high rates of metabolism in mammals (and birds) mean that relatively large amounts of heat are produced as a by-product. But you'll appreciate that this heat is not wasted; it's used in these animals to warm the body. The fact that a mammal (or bird) keeps its body temperature at a high level (37 °C - degrees Celsius - for humans) ensures that metabolism proceeds at a high and efficient rate, allowing the sustained and elevated pace of life typical of mammals. Animals that depend on internally generated heat (i.e. metabolic heat) to maintain their body temperature are called endotherms. By contrast, ectotherms have a body temperature influenced most sharply by heat from external sources, i.e. from the immediate environment.
Suppose an ectotherm - say a small lizard - is placed in a small container in which the air temperature can be varied (a useful alternative term for the temperature 'outside' is ambient temperature, Ta). The temperature of the animal's body (Tb) can be measured while we change Ta, via a miniature temperature-measuring probe into the animal's cloaca (the common opening for the gut and the urinogenital system). We could start with a comfortable ambient temperature (comfortable for a lizard, that is), say about 38 °C, record the animal's body temperature, lower the ambient temperature to 30 °C and then measure the body temperature once more, after allowing the animal to settle down over a couple of hours in the changed conditions.
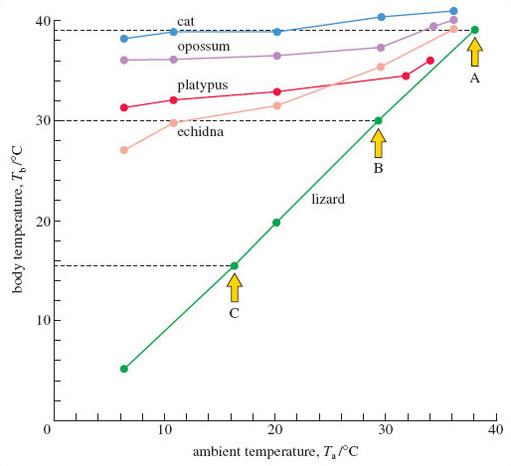
Figure 2 shows the type of results we'd get. Concentrate for the moment on the data for the lizard. In this graph, the line labelled 'ambient temperature' is referred to as the horizontal axis; that labelled 'body temperature' is the vertical axis. For a particular Ta, say 38 °C, there's a corresponding measure of the lizard's Tb - in this instance, it was 39 °C, which you can check by following the horizontal dashed line across to the vertical axis. The plotted value is identified by arrow A on Figure 2. At a Ta of 30 °C, the lizard's Tb was identical, 30 °C, as plotted on Figure 2 and identified by arrow B. For a Ta of 16 °C, the value of Tb was 15.5 °C; this point (shown by arrow C) is plotted in an appropriate position between the 10 °C and 20 °C points marked on the axes - working out where to place the point is a lot easier to do if you use graph paper. When the full range of 'lizard' values is plotted, a line is drawn between them - the lowest plotted values of Ta and Tb are 6 °C and 5 °C respectively.
The same procedure can be repeated for a placental mammal, such as the cat. Look at the cat data in Figure 2 and check that you follow the plotted points; for example, at a Ta of 10 °C, Tb is 39 °C. Joining all the plotted points gives a more flattened but somewhat 'jagged' line over the measured range of ambient temperatures.
Question 7
What's the most striking difference between the way in which the cat and the lizard respond to changes of ambient temperature?
Answer
The body temperature of the cat stays much the same when the ambient temperature falls. But in the lizard, the body temperature falls as ambient temperature drops.
So, the endothermic cat is able to maintain its body temperature - at a high and constant temperature (of about 39 °C), whereas in these conditions, the lizard has seemingly little option but to allow its body temperature to conform to that of its environment. As is typical of mammals, the cat is regulating its body temperature - more technically it is demonstrating thermoregulation. On this evidence, the lizard is a conformer - body and ambient temperature change in step. But an ability to thermoregulate is not unique to mammals; birds do so very proficiently and even some fish (such the tuna mentioned earlier) and a few insects do so, in a rather elementary fashion. You'll notice from Figure 2 that monotremes (and the marsupial representative - the opossum) are also able to thermoregulate, though less precisely than the cat.
But now suppose the body temperature of the ectotherm is recorded under sunny natural conditions, with the animal able to display its normal behaviour. Many lizards then have a body temperature considerably above that of the surrounding air, because of an ability I mentioned before - they are able to bask in the sunshine. A modest-sized lizard - for example the common European lizard - in sunny conditions, can warm from 15 °C (close to the recorded air temperature) to 25 °C in about five minutes by basking side-on to the sun. By shuttling into the sun and back into the shade, this lizard can maintain a relatively even body temperature close to 30 °C, at least during the sunnier months of the year and during the day (Tb falls at night or on sunless days). So here thermoregulation is achievable by behavioural means.
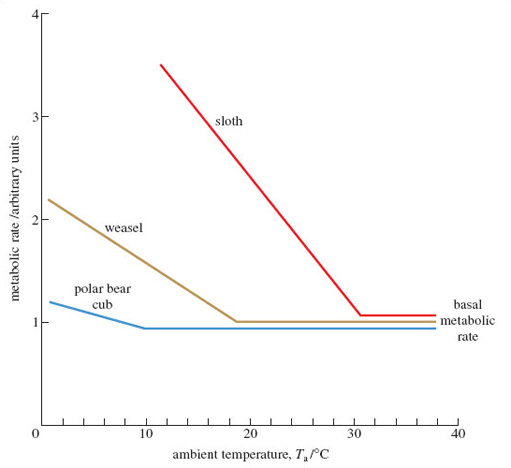
By contrast, in mammals (and birds) thermoregulation can be achieved largely by physiological means - by internal adjustments within the body's tissues and organs. Adjustments in metabolic rate are an especially important part of the process. Figure 3 helps to explain what's involved; for the moment, concentrate on the line labelled 'weasel' - which, for convenience, we can think of as a typical temperate mammal. Here, the vertical axis denotes metabolic rate; this quantity can be measured in a variety of ways - often by measuring how much heat is generated by the animal in resting conditions. Like all such scientific measurements, agreed units would normally be used to express such data, but explaining the 'what and why' we'd use here isn't crucial to our argument, so the vertical axis is labelled simply 'arbitrary units'. The horizontal axis is Ta, just as in Figure 2.
Question 8
What's striking about the metabolic rate for the weasel between the ambient temperatures of 38 °C and 19 °C?
Answer
It's constant, at what seems like the lowest possible value. Between these temperatures, there would be no change, therefore, in the amount of heat produced as a by-product of metabolism.
This minimum rate of metabolism is termed the basal metabolic rate (BMR); it would be the 'at rest' values recorded for the mammals in Table 1.
Question 9
Describe what happens when the ambient temperature experienced by the weasel falls below 19 °C.
Answer
The metabolic rate increases steadily, which means that the amount of metabolic heat produced also increases. (The fact that it's a straight line means that the metabolic rate increases by the same amount for a standard drop of ambient temperature, of say 5 or 10 °C.)
Question 10
Why might this boost in metabolic heat production be important?
Answer
When conditions become increasingly colder, the animal is likely to lose a greater amount of heat than it would in warmer temperatures. The extra heat production, therefore, might compensate for this increased loss, which helps explain why body temperature stays at a constant value in the way that we saw for the cat in Figure 2.
The type of elevation of metabolic rate we see in Figure 3 is termed a thermogenic response and is one of the most important distinguishing features of mammals (and of birds too). The type and scale of the thermogenic response varies; I can use the axes on Figure 3 to show the type of plot obtained from comparable measurements on mammals living in very different environments. When the metabolic rate in a tropical mammal like a sloth is measured against Ta, the BMR is evident over a much narrower range of Ta, and the rate of metabolism increases very sharply below about 31 °C.
Question 11
Using the correct technical terms and symbols, explain the way in which metabolism in the arctic mammal changes as ambient temperature drops.
Answer
As Ta falls, the BMR is maintained down to a Ta of about 10 °C, at which point metabolism increases comparatively slowly as Ta drops further.
The slopes of the three lines in Figure 3 differ. The steepest such slopes are typical of tropical mammals, especially those that are unlikely to encounter low temperatures. They lose heat relatively rapidly from their surface and their metabolism has to be cranked up very substantially to compensate. By contrast, mammals adjusted to very cold conditions (the polar bear cub or the arctic fox you saw in LoM) are much more proficient at conserving the heat they produce, largely because of their superb insulation. The arctic fox can maintain its BMR down to a Ta as low as minus 30 °C, i.e. −30 °C; the tropical forest-dwelling sloth is obliged to raise its metabolism at any Ta below 30 °C.
How then do mammals raise their metabolic rate in the ways shown in Figure 3? Some behavioural tactics will be familiar to you. Increasing activity - the stamping of feet - reflects an increase in voluntary muscular activity; since the muscles are working harder, they produce more metabolic heat.
Another part of the explanation is shivering, where opposing sets of muscles contract in a high frequency mode. Rather than movement resulting - the usual effect of muscle contraction - these 'inefficient' tiny contractions produce large amounts of heat. Shivering can be temporarily blocked by drugs (notably curare, which prevents muscle contraction) but when such drugs are applied, they don't entirely remove the ability that animals have to increase metabolism in the cold. This suggests that there's another component to responses of the type evident in Figure 3. It's now known that many mammalian tissues (such as the liver of species used to living the cold) have the capacity to raise their metabolism and burn off more fuel in the mitochondria. If you work through the other units in the series, you will learn about a range of circumstances when 'turning on' heat production in this way is especially advantageous. But what's striking at this stage is that mammals well-suited to life in the cold have not just those visible adaptations 'on the surface', such as thick fur. They can display 'inner' adaptations - in their metabolism, for example - of the type that only measurements such as those in Figure 3 can reveal.
5.4 Coping with heat
Not only are there the mechanisms to generate extra heat, but there are cooling mechanisms too, of which sweating is just one example.
Activity 4
Watch 'A Winning Design' on the DVD from 30.50-34.12 and write down the behavioural responses shown by red kangaroos struggling with hot, dry conditions.
Answer
They seek shade and spread saliva onto the forelimbs.
Spreading saliva onto the forelimbs achieves much the same effect as sweating; the evaporation of the fluid requires heat that comes from the skin surface. The effect is to cool the skin and the blood flowing through adjacent vessels. The flow of blood through and near the skin (i.e. the peripheral circulation) often increases, thereby augmenting the effect.
Behaviour plays an important part in promoting heat loss amongst animals that live in hot, dry environments. As you've just seen, shade seeking is one familiar example; wallowing in cooling mud is another. Echidnas live in some of the hottest environments on Earth, with temperatures often approaching 40 °C, and appropriate behaviour is crucial to their survival. No laboratory measurements (such as those in Figure 2, for example) could do justice to the subtle and diverse ways in which these animals cope with heat or cold 'in the field'. In the past, largely on the basis of laboratory studies, echidnas were thought to lack any physiological cooling mechanism. For them, a body temperature of above 38 °C is fatal, because it is at least 6 °C above their normal body temperature [p. 16]. In hot weather, echidnas become entirely nocturnal. They spend their days in caves or burrows, where temperatures are significantly cooler than outside, so their bodies can lose heat to their surroundings. But they also rest in hollowed-out logs where the inside temperature is seldom lower than that outside. The fact that body temperatures appear to stay reasonably constant (seldom more than 35 °C) in such very hot conditions suggests that echidnas may indeed have some physiological means of adjusting temperature, which may well include increasing peripheral circulation. Remaining inactive in such conditions helps reduce the amount of heat the animal produces internally. But echidnas may go one step further and actively reduce their rate of metabolism, entering a profoundly inactive state called torpor. Much the same device is used by echidnas to cope with cold conditions [p. 16], though here their prolonged inactivity (two to three weeks) and their highly suppressed metabolism (20% of the normal resting value) is more like true hibernation, with Tb as low as 5 °C.
These recent findings have come from close observation in the field and, if confirmed by other scientists, they would be further evidence that echidnas are not at all primitive and physiologically inept; they have sophisticated control mechanisms in place. Much the same is true of marsupials, though they generally have a lower Tb and BMR than do placental mammals. But far from being a primitive feature, this subtle metabolic adaptation may reduce the food requirement and ensure that food reserves last for longer during adverse conditions.
6 Thermoregulation and mammalian fur
A coat of profuse mammalian body hair is commonly called fur. Fur provides insulation, which is a property that one first thinks of as useful for mammals to help retain body heat. Fur is a unique and fundamental feature of mammals, though not all living species possess it.
Question 12
From your general knowledge, what characteristic is shared by those tropical mammals that have little or no fur?
Answer
In general, they are very large animals - think of the rhinoceros and the elephant. Large animals have the advantage of warming up comparatively slowly in hot conditions, so delaying dangerous rises in body temperature. But for the same reason, they often have severe problems when they need to lose heat. So, for very large tropical animals the loss of fur is likely to have conferred a significant advantage.
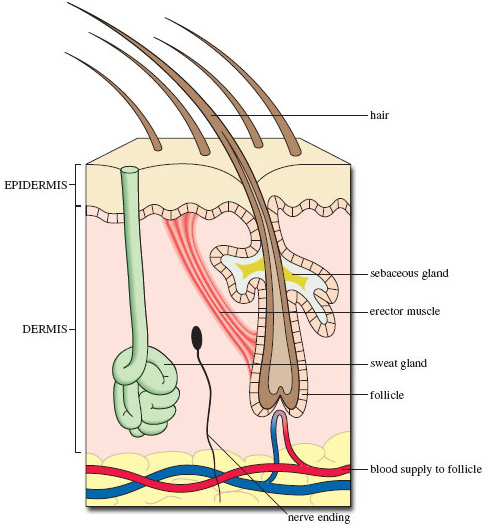
Figure 4 is a cross-section through the skin of a mammal, including a slice through a single hair. Its root is embedded deep within the inner layer of the skin, called the dermis. The sebaceous gland provides the oily secretion that helps keep the fur in good condition, while the contraction of the erector muscle makes the hair stand up; their combined action 'fluffs up' the fur, increasing its powers of insulation. In passing, note the sweat gland that is located deep within the dermis and the blood vessels that supply the base of the hair with nutrients. This point is where cell division and growth of the hair occur and the newly formed hair shaft (rich in keratin, see Section 4) pushes upwards. The part of the skin that is visible is the overlying epidermis, which is composed of cells (now dead) that were initially produced by cell division within a growing area deep within the dermis. You'll appreciate that Figure 4 is a generalised view - different mammals show particular characteristics of skin structure. For example, the skin of the echidna would look very different from that shown in Figure 4, with a mix of 'normal' coarse hair - the length of which varies according to location and therefore climate - and large single 'hairs', mentioned earlier [p. 15], which are so well reinforced with keratin that they more closely resemble our own nails in hardness and chemical make-up.
The insulation provided by fur comes not from the hair itself, but largely from the layer of air trapped within the fur. Air is a very effective insulator, which is the same as saying it's a poor conductor, i.e. it has a very limited ability to convey heat away from a warm surface, such as the epidermis. Calculations reveal that if a layer of still air of about five centimetres (i.e. 5 cm) could be held in place close to the skin, it would provide the same level of insulation as would the impressively dense winter coat of the arctic fox.
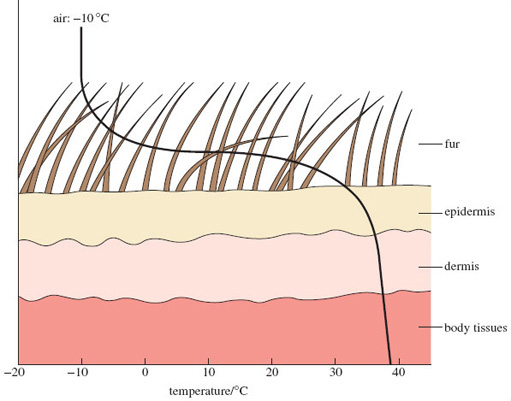
Figure 5 is a plot of the temperatures at sites within and beyond the skin surface when a well-insulated polar mammal (a wolf or an arctic fox) is exposed to an air temperature of −10 °C. Near the tips of the fur, the temperature is −10 °C but the surface of the epidermis is close to 30 °C. So a temperature difference (or gradient) of about 40 °C can be supported at either end of the 6-cm-thick fur layer, due to the excellent insulating properties of the air contained within the long, fine and densely packed fur. You can see just how little insulation is offered by the epidermis and dermis; however, the picture is different in the polar bear, which has coarse outer hair and dense underhair and relies for insulation more on blubber underlying the skin - an arrangement better suited to its amphibious existence.
You'll appreciate by now that mammals that show a very small increase in metabolic rate as outside temperatures fall (see Figure 3) do so largely because of very effective insulation. Conductance is the name given to the flow of heat from one material to another, say from the skin to the outside air. An effective coat of fur ensures a very low level of conductance of heat; the more flimsy coat of a tropical animal provides a higher conductance. So, the slope of the lines in Figure 3 is a measure of conductance - the steeper the slope, the greater the conductance.
As LoM and the TV programme have suggested, the combination of endothermy and the ability to conserve the large amounts of body heat generated has contributed enormously to the success of mammals. What is remarkable is that fur has proved sufficiently versatile during the evolution of mammals to make physiological sense for the amphibious platypus, the cold-adapted arctic fox and the sun-baked red kangaroo alike.
7 What's special about placental mammals?
As a pointer to the later units in this series, it's helpful to end this course (as LoM and the TV programme do) by stressing the differences between marsupials and the placental mammals - the mammals that provide the focus of the remainder of the units in this series. To do so, we return to the topic of mammalian reproduction.
In each mammalian type, pregnancy begins as the blastocyst becomes embedded in the tissues of the womb. In both marsupial and placental mammals, the developing tissues of the mother's womb and the embryo lie in very close contact, such that 'oxygen, carbon dioxide, nutrients and waste materials' [p. 36] can transfer between mother and offspring. In marsupials, this contact is loose and seldom any more than a very rudimentary 'placenta of sorts'. But in some marsupials, the contact between the two sets of tissues becomes more elaborate; in the koala, for example, the structure is considered by most experts to be a true placenta, albeit short-lived. The fact that a functional placenta evolved in at least a couple of marsupial species is a striking example of parallel evolution - the independent development of a structure in two distinct groups. As LoM explains [p. 37], marsupials seem to be descended from a different ancestral group than placental mammals; they can't be thought of as in any sense intermediate. It is indeed the case that there are 'different, independent solutions to the problem of rearing young' [p. 37] but sometimes, as with the marsupial 'placenta of sorts', the solutions have much in common.
The terms 'marsupial' and 'placental' were established in the late 18th century when mammals were first classified. 'Marsupial' is derived from the Latin word marsupium, meaning pocket. As you know, this feature is conspicuous in kangaroos and wallabies but is not present in all marsupials. Then the need for renaming was even more evident once marsupials were found to briefly form a simple placenta. So the terms Eutheria (= placental mammals), Metatheria (= marsupials) and Prototheria (= monotremes) were proposed instead. You may well come across these terms yourself in your own reading, but I'll be using the original and more familiar terminology in the units in this series.
What are the apparent benefits that the development of a more elaborate placenta can bring? In placental mammals, close and extensive contact of maternal and embryonic tissue reaches its peak; in the human placenta there is an estimated 40-50 kilometres of such fine-scale contact! And yet the bloods of mother and offspring never fully mix - to do so would risk the mother's rejection of the embryo as a 'foreign' object. The transfer between the two blood systems is so extensive that the needs of the growing embryo can be met for a prolonged period. The net benefit, as DA points out, is that the young of placental mammals are often born relatively mature after a prolonged pregnancy, with all the attendant benefits to their early wellbeing that size can bring. The efficiency of the mammalian placenta is another factor that helps explain the group's biological success - an evolutionary flowering that the following units illustrate to full effect.
The learning outcomes listed in the introduction to this course tell you what you should be able to do as a result of your study. You do not need to be able to do all these things from memory but you should be able to do them with the help of the text. If you have time, then you can consolidate your understanding by making some notes about each of the learning outcomes in your notebook. For example, the first one says: 'explain the distinctive biological features of monotremes.' My notes on that would say:
Monotremes (subclass Prototheria) - echidna, duck-billed platypus - lay eggs (unlike other mammals). Immature young hatch in about 10 days using egg-tooth to break soft shell, then fed on milk like other mammals.
You might like to try this technique for the other learning outcomes. It would also be useful to read through the earlier work in your notebook. Then, if you are working through all the units in this series, you are ready to begin S182_2 Studying mammals: the insect hunters.
Conclusion
This free course provided an introduction to studying Environment & Development. It took you through a series of exercises designed to develop your approach to study and learning at a distance, and helped to improve your confidence as an independent learner.
Acknowledgements
Except for third party materials and otherwise stated (see terms and conditions), this content is made available under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 Licence
Course image: tontantravel in Flickr made available under Creative Commons Attribution-NonCommercial-ShareAlike 2.0 Licence.
Grateful acknowledgement is made to the following sources for permission to reproduce material in this course:
The content acknowledged below is Proprietary and is used under licence.
Figure 4 Clutton-Brock, J. (2002) Mammals, Dorling Kindersley Limited, a Penguin Company;
Figure 5 Willmer, P. et al. (2000) Environmental Physiology of Animals, Blackwell Publishers Ltd.
Don't miss out:
If reading this text has inspired you to learn more, you may be interested in joining the millions of people who discover our free learning resources and qualifications by visiting The Open University - www.open.edu/ openlearn/ free-courses