Studying mammals: Return to the water
Use 'Print preview' to check the number of pages and printer settings.
Print functionality varies between browsers.
Printable page generated Tuesday, 9 June 2026, 10:35 AM
Studying mammals: Return to the water
Introduction
The versatility of mammals is a central theme of the 'Studying mammals' series of units, but surely no environment has tested that versatility as much as the rivers and oceans of the world. Mammals are essentially a terrestrial group of animals, but three major groups have independently adopted an aquatic way of life. In moving to the water, aquatic mammals have had to survive, feed and reproduce using a set of biological characteristics that evolved in association with life on land. This course will explore how these characteristics have provided challenges, and opportunities, for mammals that spend some or all of their time in the water.
This is the seventh in a series of units about studying mammals. To get the most from these units, you will need access to a copy of The Life of Mammals (2002) by David Attenborough, BBC Books (ISBN 0563534230), and The Life of Mammals (2002) on DVD, which contains the associated series of ten BBC TV programmes. OpenLearn course S182_8 Studying mammals: life in the trees contains samples from the DVD set. You should begin each course by watching the relevant TV programme on the DVD and reading the corresponding chapter in The Life of Mammals. You will be asked to rewatch specific sequences from the programme as you work through the course.
This OpenLearn course provides a sample of level 1 study in Environment & Development
Learning outcomes
After studying this course, you should be able to:
contrast the physical properties of air and water and describe implications of such differences for aquatic mammals
give examples of the adaptations displayed by aquatic mammals that enable them to hold their breath while submerged for relatively long periods
describe some of the biological differences between pinnipeds, sirenians and cetaceans
discuss the importance of communication by sound in aquatic mammals, describe the role of blubber and explain countercurrent heat exchange
explain the diving response and its significance in natural dives.
1 The pinnipeds, sirenians and cetaceans
As you work through this course you will come across boxes, like this one, which give you advice about the study skills that you will be developing as you progress through the course. To avoid breaking up the flow of the text, they will usually appear at the start or end of the sections.
As well as the course text, you will be using The Life of Mammals book (LoM) and related The Life of Mammals DVDs, as described in the introduction to this course. Before you go any further, watch 'Return to the Water' on the DVD and read LoM Chapter 7. Unless stated otherwise, all the page references you encounter in this course will be to LoM.
The versatility of mammals is a central theme of this series, but surely no environment has tested that versatility as much as the rivers and oceans of the world. Mammals are essentially a terrestrial group of animals: living mammals are descended from shrew-sized ancestors that evolved on land, and most mammals are still land-based animals.
For the first two-thirds of the evolutionary history of the mammals, from about 210 to 65 million years ago, dinosaurs dominated the plains and forests. But the dinosaurs conquered more than just the land: the seas and estuaries were home to other large aquatic reptiles, including ichthyosaurs and plesiosaurs. These animals disappeared at the same time as their terrestrial cousins - and their demise opened up new habitats and new sources of food that the mammals were quick to exploit.
As author David Attenborough (DA) notes in LoM Chapter 7, three major groups of mammals have independently adopted an aquatic way of life, the suborder Pinnipedia (part of the order Carnivora), the order Sirenia and the order Cetacea. These groups are the focus of our attention in this course. I'll have less to say about the smaller lineages that have become aquatic to some extent, such as the river and sea otters [pp. 185-188] that were one of the highlights of the TV programme 'Return to the Water'.
The suborder Pinnipedia
The name of this suborder comes from a Latin word meaning 'wing-footed', which refers to the modification of limbs into flippers. Most pinnipeds move onto the land from time to time to rest and bask in the sun, and in the breeding season they gather, often in large numbers, on beaches or flat areas of sea-ice to give birth. There are three main families: the eared seals, which include fur seals and sea-lions; the earless or true seals; and the walrus. Remember that these mammals are carnivores; in view of their continuing links to land, they are sometimes aptly called 'amphibious carnivores'.
The other two groups, the sirenians and cetaceans, are fully committed to a life in the water.
The order Sirenia
There are four living members of the order Sirenia: three species of manatees and the dugong. These animals are descended from ancestors that also gave rise to the elephants, and they are the only herbivorous, completely aquatic mammals. As DA points out [p. 200], the order is named after the Sirens of classical mythology, sea-nymphs who lured sailors to their death. Until relatively recently, there was a fifth species: Steller's seacow. This animal lived mainly on kelp and had no teeth, though it did have rough plates on its palate. Hunting and the decline of the kelp beds caused a drastic fall in its numbers, and it has the dubious record of the shortest period of time between discovery by western science (1741) and extinction (1768).
The order Cetacea (dolphins, porpoises and whales)
The cetaceans get their name from the Greek word for a whale (ketos). The order is divided into two suborders: the Odontoceti, or toothed whales, which include the sperm whale, killer whale, dolphins and porpoises; and the Mysticeti, or baleen whales, which include the blue whale and humpback whale, and are named for the brush-like plates that they use to filter krill from the water, as you saw in the TV programme at 35.40.
In moving to the water, aquatic mammals have had to survive, feed and reproduce using a set of biological characteristics that evolved in association with life on land. This course will explore how these characteristics have provided challenges, and opportunities, for mammals that spend some or all of their time in the water. I shall examine some of the important differences between a life in air and a life in water, and describe how the bodies and behaviour of aquatic mammals have changed in ways better suited to living and hunting in an aquatic environment. In particular, I shall explore the so-called 'diving response' (Section 3), and describe some of the problems involved in finding out how the animals behave on their dives. Finally, in Section 4, I shall look at what is now known about the evolution of the mammals that have made the most complete transition from the land to the water - the whales. So, in the early part of this course I'll be revisiting some of the topics introduced by DA in LoM and the TV programme; my aim here is to consolidate what you've learnt already. Later on, we move into uncharted waters to cover new topics that build on what's gone before. This course also gives you an opportunity to read two intriguing articles on diving mammals, one written by a science journalist and the other by a team of research scientists.
You have just met the scientific names of some mammalian groups (orders and suborders) with explanations of how the words have been derived. Many scientific names have Latin or Greek roots and with a good dictionary (and maybe a web search), you may be able to unravel some of these words for yourself. For example, the Weddell seal which you will meet later in this course has the scientific name Leptonychotes weddelli (remember that scientific names are always printed in italics; if you write them by hand, you should underline the name). My dictionary gives, at the end of the definition of the word 'lepton', the Greek word leptos meaning 'slender' or 'fine' and at the end of the definition of 'onyx', it gives onyx as a Greek word meaning 'nail' or 'claw'. So the name of the genus, Leptonychotes refers to the narrow claws on the flippers of the seal (actually, the hind flippers in this case). The species name weddelli is given in honour of Captain James Weddell, who described and illustrated the seal in his writings about an Antarctic voyage to catch seals in the 1820s. You might like to try to unravel an example for yourself - try the Pacific white-sided dolphin Lagenorhynchus obliquidens. (Hint: use a comprehensive dictionary to look up 'lagena', 'rhyncho-' and 'oblique'.)
2 Living in the water
2.1 Land versus water
Mammals share a number of biological characteristics that mark them out as members of the class Mammalia. Many of these are adaptations to a life on land. For example:
Mammals give birth to young at a relatively advanced stage of development and feed their young on milk.
Most mammals have hair, or fur, covering part or all of the body.
Mammals have a high metabolic rate and maintain a relatively high and constant body temperature, irrespective of the temperature of their environment.
Most land mammals walk or run with a leg at each corner - a limb pattern they share with most other land vertebrates, including reptiles (except snakes and legless lizards) and most amphibians.
Mammals breathe air and have a four-chambered heart, allowing them to split their circulation into a system for passing blood through the lungs and a system for transporting oxygen-rich blood around the body.
Almost all mammals can swim to some extent - even bats - and many make their way into the water from time to time for one reason or another.
Activity 1
Watch the TV programme from 00.35-08.56, which shows an elephant swimming in the sea, a desman hunting in a river and sea otters off the Californian coast. As you watch, make notes about what might tempt a land animal into the water in the first place, and the main challenges it would face in the new environment. Note how the desman and the sea otters have overcome some of these difficulties, and the devices that allow another mammal, David Attenborough, to join the otters in the water.
Before you watch, you might like to know a little more about the evidence for DA's passing remark that elephants might have adopted a semi-aquatic way of life at some point in their evolutionary history. The lungs of modern elephants are surrounded by loose connective tissue, not the bag-like 'pleural cavity' found in other land mammals, and this allows them to resist some of the damaging effects of water pressure. In addition, the kidneys of elephant fetuses possess features that are common in aquatic mammals but not land mammals. If the theory is true, the ancestors of today's elephants must have moved into the water and then returned to the land, while their sirenian cousins went on to become fully aquatic.
Answer
The obvious attraction for the desman and the otters is food, but other mammals might take to the water to escape from predators, pests or extremes of temperature. (The elephant is now a land mammal, of course, but it enters the water to drink or bathe and it can swim across open water using its trunk as a snorkel.)
The main challenges for the desman appear to be holding its breath long enough to find and catch its food, moving about in the strong current and staying warm in the water. The sea otter also holds its breath while it uses webbed feet and a rudder-like tail to search for food in shallow coastal waters, and its thick fur keeps it warm enough to live in Alaskan seas. DA can join the otters, thanks to a snorkel (for breathing), fins (for moving about), a wetsuit (for staying warm) and a mask (for seeing in the water).
Watching the TV programme you might have thought of other examples of aquatic mammals: both the yapok (a marsupial) and the platypus (a monotreme) are very much at home in the water, despite the demands of that environment. The challenges that face the platypus, desman, sea otter and DA are common to all aquatic mammals. They are the result of matching their mammalian characteristics against a number of differences in the physical properties of air and water, the most important of which are listed below.
Water contains less oxygen than air does.
Water is denser and more viscous than air.
Water has a higher specific heat and a higher thermal conductivity than air. (I'll explain these terms later.)
Light is much more readily absorbed by water than by air and visibility in water is often poor because suspended particles make it turbid.
Sound travels faster and further in water than it does in air.
2.2 Breathing air
A swimming elephant can breathe by holding the end of its trunk out of the water, but if it tried to find its food under the surface, like the desman, it would have to hold its breath. Neither the mammalian lung nor the skin can extract enough oxygen from water to sustain life, so aquatic mammals must come to the surface at intervals to breathe; and all of them - pinnipeds, sirenians and cetaceans - drown if they are prevented from doing so for prolonged periods.
Lungs form 7% of the body mass of a human being, but no more than 3% of the body mass of most cetaceans - so lung size alone cannot account for their diving abilities. Part of the secret is that aquatic mammals use their lungs more efficiently: they extract more oxygen from the air they take in, and change a higher proportion of the air within their lungs with every breath. For example, we take in about a litre of air with each breath, despite the fact that our lungs can hold over four litres; a fin whale takes in 3000 times that amount in just under two seconds.
It is the ability to fill and empty their lungs so quickly that produces the 'blow' of the larger whales - something so distinctive that it can be used to identify species, even individual whales, far out at sea. The animals pay a price for this efficiency, however. We can obtain extra oxygen by taking deeper breaths; a whale can only breathe more frequently. Whaling ships used to take full advantage of this fact, chasing whales until the exhausted animals were forced to spend more and more time at the surface.
The need to breathe air means that sirenians and cetaceans have to be able to sleep without drowning, as these animals never leave the water. The blow-hole in cetaceans seems to be under voluntary control, which suggests that these animals have to be 'awake' to breathe. Whales and dolphins rest in the water from time to time at or close to the surface, and animals in social groups spend periods swimming slowly side by side ('echelon swimming'). This paired swimming mimics the behaviour of a calf with its mother and is probably associated with rest. It is likely that at these times the animals use a form of 'cat-napping', in which one half of the brain rests while the other remains active.
2.3 Moving about
Water is more viscous than air, so it can take more effort to move through water (try running in a swimming pool). Friction between the body and the water causes turbulence, which holds a swimmer back, and the faster the swimmer tries to move, the greater the turbulence. One way of avoiding the problem is to leave the water for short periods and travel through air, and some of the smaller pinnipeds and cetaceans resort to 'porpoising', leaping from the water for short periods when they need to move as quickly as possible. But to catch their food, these animals have to be swift and agile in the water.
Pinnipeds breed on land (or ice) so they have to be able to move about on land as well. They have four flippers arranged in the typical mammalian limb pattern: two at the front and two at the back. In true seals, the hind flippers project behind the body and provide extra propulsion in association with a sideways swimming motion [p. 188]. The shorter front flippers are used for steering in the water and for pulling the body along on land. Fur seals and sea-lions use longer front flippers to 'fly' through the water; the hind flippers are mainly for steering and most of the power comes from the body muscles and spine. On land, however, the hind flippers can be brought forward beneath the body, allowing the animal to lift itself up and 'walk'.
As you know from LoM, sirenians and cetaceans are fully aquatic and never leave the water. They swim in a rather different manner, using an up and down motion of a tail with horizontal flukes. The front fins help with steering and keep the animal stable in the water. This vertical swimming motion is another legacy of the animals' terrestrial ancestry: imagine how the spine of a sprinting cheetah (or a galloping horse) flexes to allow the animal to increase its stride length.
The muscles of aquatic mammals are no stronger per unit mass than the muscles of terrestrial mammals, so a dolphin's top swimming speed of around 50 km h−1 is not the result of muscle strength alone. As DA explains, most aquatic mammals are streamlined to some extent to help them slip through the water: a smooth, cigar-shaped body with few external projections reduces water resistance and the drag created by turbulence. Streamlining is so important for aquatic animals that many fish, aquatic reptiles such as the ichthyosaur, and aquatic mammals share a common body shape - another example of convergent evolution (see Figure 1). The streamlining of cetaceans is made even more effective by a special property of their skin. The outer layer is soft and spongy, and reduces turbulence at the skin-water interface by expanding and contracting in response to the flow of water.
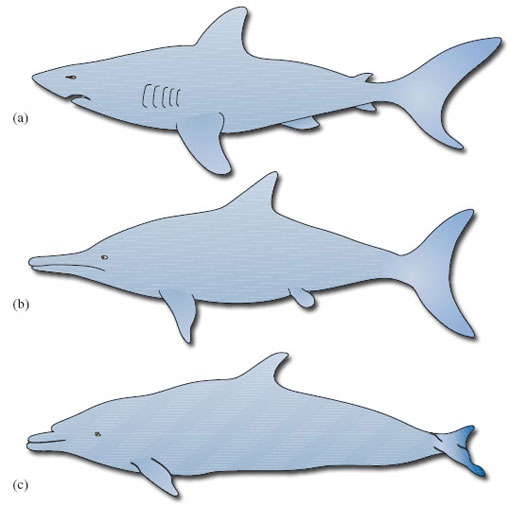
These adaptations are so effective that it is now believed that cetacean swimming is pretty near as cost-effective (in terms of metabolic energy expended for distance travelled) as the running of land animals. Swimming at the surface is more expensive, however, as is the swimming of semi-aquatic mammals such as otters. It appears that economic running and swimming are difficult to combine in the same animal.
2.4 Staying warm …
In this section, you will meet the term 'thermal conductivity' and you will be asked to accept that it is 'a measure of how readily heat flows from a particular material'. You may be uncomfortable about the lack of detailed explanation of how it is measured and of actual values and units. However, at all levels in science, there are times when we must accept a less-than-complete understanding and be prepared either to accept this, or to search out more details for ourselves. In particular, in a course of this level, there will be aspects that have to be taken on trust for now.
Water has a higher specific heat than air: that is, it takes more energy to raise the temperature of a particular volume of water than it does to raise the temperature of the same volume of air by the same amount. One consequence of this is that the temperature of surface waters tend to fluctuate within a smaller range than air temperature: from −1.9 °C (the freezing temperature of seawater) near the poles to an extreme of about 30 °C in the tropics. But deep water is always cold, whatever the location in the ocean; below about 1000 m, the temperature is uniformly cold, about 3-4 °C. The risks of heat loss from an aquatic mammal are all the greater because water has a relatively high thermal conductivity [p. 184], so a warmer object loses heat fairly quickly when immersed. (Think of thermal conductivity simply as a measure of how readily heat flows from a particular material.) Marine mammals, like many of the larger land mammals, maintain their body temperature somewhere between 35 °C and 38 °C, so they all face the problem of staying warm in a colder environment.
To put this into some sort of context, consider the Weddell seal of Antarctica. It maintains an internal temperature of 37 °C, despite water temperatures of around 0 °C and air temperatures of 0 °C to −50 °C (with a wind chill down to −100 °C or so). The newborn pup is protected from the cold by a dense layer of fur called 'lanugo'. In the first month of so of life, as it prepares to go to sea, the pup replaces its lanugo with the shorter, sparser hair of the adult seal and develops a thick layer of blubber.
Question 1
Can you think of a likely explanation for this change?
Answer
As DA points out, blubber provides better insulation in the water (and the smoother coat of the adult improves its streamlining).
A fur coat traps a layer of warm air next to the body and is a fairly good insulator in air and water, but it has its limitations. It requires a lot of maintenance, and water pressure squeezes out the air if the animal travels too far below the surface. (It is the still air trapped by fur that provides the insulation; a trapped layer of water in fur would provide some insulation, but not nearly as much as that offered by air.) Otters and fur seals make relatively modest dives [pp. 185-188], but many cetaceans and true seals, like the Weddell seal, hunt in deeper, colder waters.
As DA points out, animals that dive to any great depth stay warm using blubber, a thick layer of fatty tissue that extends far beneath the skin. In air, blubber provides less insulation than an equal thickness of fur, but it retains much of its effectiveness in water and the extra bulk is less important in the more supportive medium. (You'll appreciate the fact that the relatively high density of the surrounding water helps 'buoy up' aquatic animals, such that diving mammals weigh relatively little in water.)
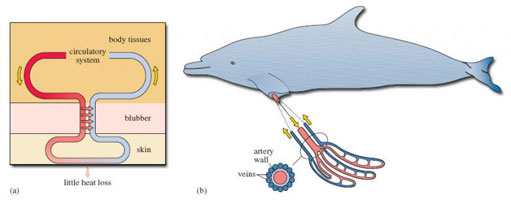
Those parts of the animal not covered in blubber - the limbs and parts of the head - have another device for retaining heat: a countercurrent heat exchange system. In Figure 2a, the outgoing (i.e. arterial) blood is shown to the left, on its way to the skin surface. But en route, such vessels run very close to a network of veins that are bringing cool, venous blood back into the body (shown to the right in Figure 2a). Given the closeness of the (warm) outgoing and (cooler) incoming bloods, heat (which would otherwise be lost through the skin) is taken up by the cooler returning blood and carried back inside the animal. The red arrows in Figure 2a show the direction of heat flow; there is comparatively little heat loss from the skin. If you think about it, the system depends on outgoing and incoming blood flowing in opposite directions, which is why it is called a countercurrent heat exchanger. (If the two blood flows were in the same direction, there would cease to be any heat transfer once the blood temperatures had become identical.)
The system is shown to best effect in the flippers (and tails) of diving mammals, which have to be thin, and therefore free of insulating blubber, if they are to function effectively. Efficient heat exchange at such sites is essential if heat loss is to be avoided. Figure 2b shows the close juxtaposition of the arterial and venous bloods, with the central artery and surrounding veins in cross-section.
Fur and blubber are so effective as insulation that some aquatic mammals can find themselves with another problem: losing excess heat after exercise, especially in sunny conditions. Blubber is full of small blood vessels which dilate to bring warm blood close to the skin, allowing a certain amount of active temperature regulation. The countercurrent system has its own control mechanism. A rise in blood pressure with activity causes the arterial walls to expand, which closes the network of veins bringing cool blood back inside. The warm blood then passes straight through to the skin, excess heat is lost to the external environment, and the cool blood has to find another (non-countercurrent) route back to the heart.
2.5 … and getting larger
Size offers a number of biological advantages, including some protection from predation, but it can also help in reducing heat loss. A large mammal has a large body mass (generating heat) relative to its surface area (losing heat). But there will be constraints on any evolutionary trend of increasing body mass in some aquatic mammals. Seals, sea-lions and toothed whales have to remain agile enough to chase their prey, and the pinnipeds as a whole have to be able to move about on land, but they are all still fairly large animals. Manatees live in relatively warm waters. Their metabolic rate is comparatively low and they normally expend relatively little energy on keeping their body temperature constant. Their large size helps conserve body heat but they are very vulnerable to cold water, below about 15 °C.
The baleen whales, however, feed by filtering fish or invertebrates as they cruise the colder oceans of the world. As their prey is not chased, and their bodies are well supported by the density of the water, over the many millions of years of their evolution they have grown to immense sizes. The blue whale is the largest animal that has ever lived, weighing in at up to 150 tonnes (an average bull African elephant is around six tonnes). DA's enthusiastic reactions to the surfacing of a blue whale and his leisurely walk within a blue whale skeleton as it gradually takes on a life-like form (in the TV programme at 36.52) both give a powerful sense of the animal's impressive bulk.
2.6 Senses and communication …
Glance down at the second paragraph of this section, where you will find a sentence about the speed at which eyes adapt from bright light to poor light, and the statement that this process takes 20 minutes for the human eye. With your developing sense of scientific enquiry, that might lead you to wonder how it would be possible to test this assertion. Could you design an experiment to see if it does take 20 minutes for the human eye to adapt fully to poor light?
Light travels only a few hundred metres through clear water, so sight is of limited use to animals hunting in deeper waters.
Seals use their eyes and sensory whiskers to search for food in relatively shallow water. Elephant seals feed at depths of 300-700 m and have eyes that adapt to poor light faster than those of any mammal tested (some six minutes compared with 20 minutes for the human eye). Despite this capacity, vision is probably of little use at the bottom of their feeding range and they must rely on sensitive whiskers and hearing. In fact, the whiskers of seals and sea-lions are so important that a special blood supply at their bases keeps them warm and flexible even in the deepest, coldest waters.
Eyesight is certainly of some use in making sense of the immediate environment, and sirenians need little else to find their food in shallow, clear waters. But you'll appreciate that to hunt at depth, as many toothed whales do, communication using sound is generally a better option. Sound travels five times faster in water than it does in air, and the toothed whales use a range of high-frequency squeaks, whistles and moans to communicate with each other and coordinate social behaviour, of the type that you saw very dramatically in the coordinated driving of fish by a small pod of bottlenose dolphins off the coast of South Carolina (from 32.50-35.38 in the TV programme).
You'll also be aware that dolphins (and many other toothed whales) use a sophisticated echolocation system - 'seeing with sound' - to make sense of their environment and detect prey.
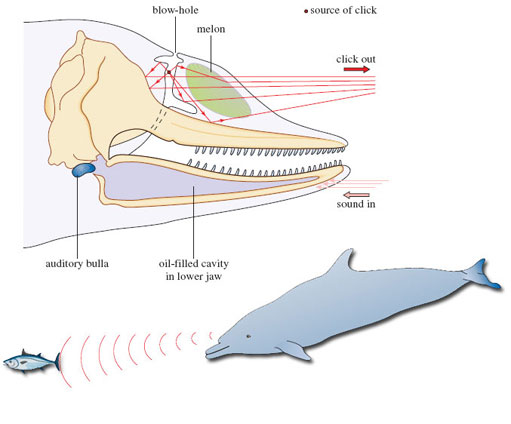
Dolphins produce a range of high-frequency clicks using air passages around the blow-hole. These clicks are focused by the front of the skull, which is shaped like a satellite dish, and pass out into the water through a waxy structure, the 'melon', which acts as an acoustic lens. In this way, the animal can focus and direct the stream of sound. The returning pulses are picked up by oil-filled cavities in the lower jaw and pass through to the inner ear, which is enclosed in a bone casing called the auditory bulla (Figure 3). This bone casing itself is surrounded by a mucus foam to isolate it from other vibrations. The whole system is sensitive enough to allow the animals to determine the size, shape and distance - even the internal structure - of objects as much as 800 m away. It is also powerful enough to allow them to stun, or even kill, their prey with a well directed burst of sound.
As well as travelling faster in water, sound also travels further and with less distortion than in air. The other suborder of cetaceans - the baleen whales - utilise this property in their use of low-frequency calls produced from the vocal cords to communicate with each other across the oceans. The song of the humpback whale is perhaps the best-known example. The song differs from population to population, and individual to individual, and it develops over time as the whales imitate each other and introduce variations. The whales also exploit a special property of the oceans to make themselves heard over great distances. At a depth of around 1000 m, there is a band of water in which temperature and pressure combine to reduce the speed of sound to a minimum. The low-frequency song of the whales can spread out through this 'sound channel' for distances of up to 1000 km, with little loss of energy to the water surrounding the channel.
Recent research on harbour seals and killer whales in the northeastern Pacific illustrates the sophistication of the sound world of marine mammals. The seals largely ignore a resident population of fish-eating killer whales, which use frequent echolocation clicks and communication calls. From time to time, however, the area is visited by transient pods of mammal-eating killer whales. These animals use few vocalisations, presumably to avoid alerting the seals, but the seals have learned to recognise the sounds that they do make and take evasive action.
2.7 … and becoming more intelligent
Intelligence is a useful commodity: it can help an animal to make sense of its environment and cope with the demands of social behaviour (including courtship and competition). Hunters tend to be relatively intelligent, and otters, pinnipeds and cetaceans, for example, share a playful curiosity that is characteristic of animals that catch other animals for a living. Some especially extravagant claims have been made for the intelligence of the toothed whales, largely because these animals use communication and cooperation to maximise the efficiency of their hunting behaviour.
Activity 2
Watch the TV programme from 32.50-35.38, which shows the hunting behaviour of the bottlenose dolphins off the coast of South Carolina and the 'bait-balling' of the common dolphins in the open ocean. Also reread LoM pp. 206-208. Note down any aspects of the animals' behaviour that strike you as intelligent.
Before you watch, it might be an idea to spend a few minutes thinking about what 'intelligent' means. But be warned, it's by no means an easy concept to pin down.
Answer
I noted the following points:
Strategy and tactics: the dolphins identify a suitable target (a shoal of fish) and plan ahead (edging the shoal close to the bank or the surface of the water).
Teamwork and cooperation: working together to achieve a common goal.
The use of communication to coordinate behaviour.
The adoption of specific roles within the group (the 'spy-hopper', for example [p. 207]).
The use of judgement to select the right option at the right time.
Some of the words I noted down certainly imply intelligence - 'planning', 'communication', 'judgement' - but it is difficult to define or measure intelligence in ourselves, let alone other animals. The behaviour of the dolphins certainly seems as sophisticated as the hunting strategies of lions or African hunting dogs, for example, but is there any evidence that they are more intelligent than that?
Well, cetaceans have large brains relative to body size, with folds in the part of the brain responsible for complex functions, the cerebral cortex, that are reminiscent of those in primates. They use sound in a number of sophisticated ways and engage in subtle social interactions that may be mediated by a simple form of 'language'. Dolphins can be trained to respond to commands and perform tricks - and they can develop and vary these routines and imitate each other's behaviour. On the other hand, the large brain and the folding of the cerebral cortex may be simply a function of the size of these animals, or a consequence of the need to produce and process such complex sounds. And many other animals can be trained to perform tricks, including pigeons and other species not generally regarded as particularly bright.
One behaviour that has caused some people to question the intelligence of cetaceans is a tendency to strand themselves in large numbers on beaches and, even more puzzling, to strand themselves again if they are helped back into the water. The reasons for this phenomenon are poorly understood. It could be a response to distress calls, a form of mass panic, or the result of damage to the echolocation system caused by disease, parasites or pollution. As with so many aspects of cetacean behaviour, the honest answer is that we just don't know.
One difficulty in interpreting cetacean behaviour may be our tendency to describe and discuss intelligence in terms of the way we, as humans, interact with each other and our surroundings. The arrogance of attempting to judge the intelligence of other species by our own standards was used to comic effect by Douglas Adams in The Hitchhiker's Guide to the Galaxy.
It is an important and popular fact that things are not always what they seem. For instance, on the planet Earth, man had always assumed that he was more intelligent than the dolphins because he had achieved so much - the wheel, New York, wars and so on - whilst all the dolphins had ever done was muck about in the water having a good time. But conversely, the dolphins had always believed that they were far more intelligent than man - for precisely the same reasons.
(D. Adams (2002, first published 1979) The Hitchhiker's Guide to the Galaxy, Picador)
3 The 'diving response'
3.1 Features of the diving response
All the aquatic mammals' adaptations to life in the water - breathing, moving, staying warm and making sense of the environment - come together in their diving behaviour, and the diving abilities of marine mammals are truly awe-inspiring. The elephant seal, for example [p. 192], makes repetitive, long-duration dives and some 80-95% of its time at sea is spent submerged. Its dives are of about 20 minutes duration on average, and the intervals at the surface are seldom more than about two minutes. This pattern is repeated almost continuously for between two and eight months. Such abilities are especially useful for those seals that often travel long distances under ice from one breathing hole to another - the Arctic-dwelling ringed seal shown in the TV programme at 18.59 is one such example and its Antarctic counterpart, the Weddell seal, is another. The sperm whale routinely swims down to depths in excess of 1000 m to hunt giant squid and can stay submerged for over an hour.
The length of time a mammal can spend under water depends on the amount of oxygen it takes on board before a dive and its ability to slow the rate at which it uses that oxygen once the dive is under way. It was not until the 1930s and 1940s that biologists began to understand the diving abilities of marine mammals. Work on captive seals making simulated dives in the laboratory - mainly Weddell seals and elephant seals - identified a number of physiological mechanisms that became known as the 'diving response', detailed below. Subsequent research involving monitoring animals in their natural environment has shown that the full diving response is necessary only in extremis, when the animal is pushed close to its physiological limits.
The animal stops breathing (technically known as 'apnoea'.
The heart rate slows very markedly - a condition termed bradycardia. Many seals can reduce the rate to about four beats per minute.
The blood pumped by the heart is diverted away from peripheral tissues and muscles to the oxygen-dependent heart, brain and other essential organs. In essence, the animal turns itself into a 'heart-lung-brain' machine.
The overall metabolic rate of the diving animal falls as some tolerant tissues - the gut and kidneys, for example - are starved of oxygen. The metabolic rate also falls as the temperature of peripheral tissues - the flippers or flukes, for example - moves closer to that of the surrounding water. (In general, the warmer a tissue the greater its metabolic rate and, hence, the greater the demand for oxygen.) The countercurrent heat exchanger ensures that the blood supply to these peripheral tissues is maintained even as tissue temperature falls (Figure 2b).
The animal swims in a slow and sustained manner, gliding along to conserve oxygen.
These adaptations allow the animal to make efficient use of the oxygen carried on the dive. Two other adaptations allow the animals to increase the amount of oxygen carried, providing what some biologists have called a 'physiological scuba tank'.
As DA explains [p. 194], oxygen is stored by two proteins in the body, haemoglobin in the red blood cells and myoglobin in the muscles, and diving mammals have exceptional quantities of both substances.
In some diving mammals, muscles in the spleen contract to squeeze out more oxygen-holding red blood cells when the animal dives. The spleen is also larger, allowing it to hold and release more red blood cells.
Perhaps surprisingly, the main elements of the diving response are not confined to aquatic mammals. A similar response is found in all mammals and most other air-breathing vertebrates, including reptiles and birds. The observation that the diving response seems to be 'hard wired' into all mammals has led to some debate about its wider purpose. It may be that it has a role in other situations in which the body has to cope with reduced levels of oxygen.
Question 2
Can you think of any circumstances in which the ability to withstand oxygen starvation might be of use to a terrestrial animal?
Answer
There are a number of possible situations, including vigorous exercise or oxygen starvation during birth.
Of course, we are also mammals and we have our own diving response. The PDF reading 'Into the abyss', linked below, describes how humans have explored the limits of their mammalian diving skills to develop a new sport: freediving. Freedivers swim as deep as they can without the aid of scuba tanks. A normal, healthy human being can endure apnoea for one to two minutes and dive to a depth of 10-20 m. But in June 2000, Loïc Leferme dived to a depth of 152 m, and he and his colleagues regularly stay under the water for seven minutes or more. Freedivers can also reduce their heart rates to an amazing six beats per minute, which is not far away from the minimum of a diving seal.
Click 'View document' to open Reading 1: 'Into the abyss'
3.2 Natural dives
The physiology of the diving response can be studied in the laboratory, but investigating the behaviour of a diving mammal in its natural environment can be more of a problem. However, modern physiological techniques have made it possible to record continuously physiological variables (such as heart rate) and information on depth and position during the spontaneous dives in the wild that are part of the animal's normal behaviour. Most such findings show that the majority of an animal's dives don't approach the 'Olympic record' limits that represent maximum effort. For example, in the Weddell seal most dives were shorter than 20 minutes and there is little evidence of profound adjustments in the heart and circulatory system during this time, or of the presence of lactic acid in the blood of these animals as they surface. Lactic acid is an important indicator of the onset of anaerobic metabolism (i.e. metabolism that doesn't use oxygen). The production of lactic acid is a relatively inefficient way of releasing energy by the breakdown of simple sugars obtained from the diet. (In humans, lactic acid appears in the bloodstream during intense activity such as sprinting, when the supply of oxygen to the muscle cells isn't sufficient to allow the oxidation of sugars in the 'conventional' way.)
Question 3
What do these observations suggest about the physiological events that accompany natural dives?
Answer
They suggest that a full-scale diving response is not typical of these shorter dives. If tissues were starved of oxygen, substantial levels of lactic acid would probably be evident in the blood, as anaerobic metabolism became established.
The implication is that in shorter dives, the amounts of oxygen carried by the Weddell seal (mainly in the blood, as we've seen) are sufficient to tide the animal over, without a sustained or profound diving response. But with natural dives of 45 minutes or so, anaerobic metabolism becomes much more significant and the selective re-routing of blood (together with bradycardia) becomes much more important.
What about the animal's hunting behaviour? The Weddell seal, for example, spends most of its time hunting cod and other fish out of sight beneath the Antarctic ice.
Marine biologist Terrie Williams and her colleagues at the University of California have studied the hunting behaviour of Weddell seals by attaching recording equipment to animals captured near their feeding holes. The seals are fitted with video and audio recorders, plus monitors that note the animals' depth, speed, compass bearing and flipper strokes. The seals are then released back into the sea. The information that the scientists receive allows them to construct a three-dimensional map of the dive that is tied to specific pieces of behaviour recorded by the video camera. The seals hunt in three dimensions in low levels of light and have to find, stalk and catch their prey while holding their breath.
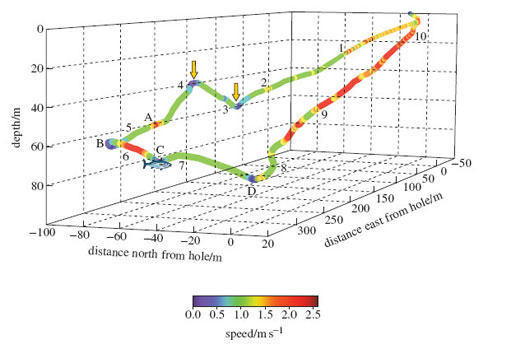
Figure 4 shows a dive by an adult female Weddell seal. The dive lasted for 10 minutes 30 seconds, and the animal travelled a total of 760 m. About mid-way through the dive, at point C, the video footage revealed that the seal attacked a large Antarctic cod, approaching from behind and below its tail. Let's look at what happened in more detail.
The seal started the dive with a few powerful strokes of its flippers. Water pressure would have quickly collapsed its lungs, making the seal denser than the surrounding water. The negatively buoyant animal could then have continued its descent with little effort. (The team have discovered that in pinnipeds and cetaceans this effortless gliding minimises the effort expended by the animal while submerged.) The seal sank slowly to a depth of 51 m (right arrow), rose slightly to a depth of 33 m (left arrow), and then started a new descent, maintaining roughly the same bearing and speed. This course was taking it in the general direction of the fish.
At point A, some 23 m from the fish, the seal used a few large flipper strokes to change direction and speed, accelerating away from the fish at an angle of 58 degrees to its previous course. It then used a looping turn (point B) to bring it back towards - but beneath - the fish. It accelerated again towards the target but slowed just before contact was made. After the attack had failed, the seal did not pursue the fish but continued its descent for a while, before turning left (point D) and proceeding quickly back to the ice hole.
Question 4
The data gathered tell us a lot about what happened but little about why. We are still left with the problem of interpreting the sequence. Try this for yourself. For example, why might the seal have moved away from the fish at point A?
Answer
Here is my interpretation (and it is difficult, if not impossible, to prove). I suspect that the initial descents and ascents on a constant bearing were a searching strategy. At point A, the seal spotted the cod a short distance away (remember the poor light). If it approached the fish head-on it would be seen quite quickly. The turns at points A and B allowed the seal to attack the fish from behind and below.
After accelerating out of the second turn, it pulled its flippers to its side and coasted towards its prey, which accounts for the deceleration recorded just before contact. The seal would be camouflaged against the darkness of the deep water and the fish would be silhouetted against the ice.
If you find this topic as interesting as I do, you have the opportunity to find out more from Reading 2, which reproduces in full the original scientific paper by Terrie Williams and her colleagues that reported these finding to the scientific world. The language used in the paper is sometimes technical and assumes some familiarity on the part of the reader with specialist terminology and the use of recording techniques. But you don't need to understand it all to get something from the paper. You'll see that more information is provided than was possible in the much abbreviated description of the research I've just presented. The reasons the research work was undertaken are outlined, and a fuller account provided of the three brief encounters - from 57.4 hours of recording! The broader significance of these findings is discussed, with an emphasis on what was new and unexpected about these observations and the potential of the technique for revealing more about the hunting behaviour of diving mammals.
Click 'View document' to open Reading 2
Part of the enjoyment of study is the opportunity to read more about a subject of particular interest, and the two papers linked with Section 3 reflect two very different types of publication available. The first reading, 'Into the abyss' from the popular science magazine New Scientist, is easy to read in that it assumes little specialist knowledge from the reader. Its style is dramatic and involving - look at the opening sentence: 'It's deathly dark, wet and you're chilled right through'. It makes you want to read on. Publications such as New Scientist or BBC Wildlife provide an excellent means of developing your interest in topics that have caught your attention in this course. But relying solely on articles written by science journalists as your source of information can cause problems, despite the many excellent articles produced, especially in the publications just named. Assertions made may not always be backed by evidence and sometimes accuracy and caution are sacrificed for the journalistic purposes of a particular angle to capture the reader's attention. And dealing with so many snippets of journalistic information may prove difficult. The over-arching concepts and ideas - the scaffolding that binds bits of information together - can be lost sight of, which is where the formal study of the subject, perhaps through OU courses, will help.
Reading 2, 'Hunting behaviour of a marine mammal beneath the Antarctic fast ice', is an example of a research paper, reprinted from the American journal Science. Such a publication is geared much more towards the interests and specialist knowledge of fellow scientists. It is included here to give you an idea of how research in this field is undertaken and the form in which it is published. The writing is much more measured and cautious in tone, describing the results in detail and stressing how the findings relate to overall knowledge of the field. For a fellow researcher in the field, the technical detail provided here, for example in footnote 6 on page 236, would be invaluable, but for most of us the details are more than we need to know! If you study more science, research papers will assume a greater importance, but at this level it's more important to know what potential benefits they offer, rather than to spend precious time struggling with the details. Most original research papers are even less easy to read than this example!
4 The evolution of whales
4.1 The rate of evolution
I now want to move away from looking at the challenges facing all aquatic mammals, to examine very briefly what we know about the evolutionary history of the cetaceans. This group has travelled furthest from its terrestrial roots and made the fullest adaptation to life in the sea.
Since mammals evolved on land, it has long seemed reasonable to suggest that the origin of whales must have involved an evolutionary transition from the land to the water. But how can we explain the fact that living whales are so well adapted to a life in the water, and so far removed in form and behaviour from most other mammals? There are two main problems. First, if evolution proceeds through the build-up of numerous small changes, it is difficult to see how the wide gap between terrestrial mammals and whales could have been bridged - even given the enormous amounts of time involved. Second, there is the problem of 'intermediate forms': that is, the nature of the animals that must have filled the gap.
In the 1930s, the American biologist George Gaylord Simpson noted something interesting about the apparent rate of evolutionary change in whales. He examined the fossils of some primitive whales, which had been dated back to the Eocene (about 50 million years ago), and compared the anatomy of these early proto-whales with the anatomy of their living descendants.
One can measure the average rate of evolution for various anatomical features in the 50 million years it took to modify [primitive] Eocene whales into fully modern forms. Let us then take that measured rate of evolution within whales and calculate how long it would have taken for Eocene whales to evolve from terrestrial ancestors. Extrapolating back, it would have taken at least 100 million years (possibly even considerably more) for the transition from terrestrial ancestor to aquatic, primitive whale descendant to have occurred - assuming, that is, that whales evolved from terrestrial ancestors at the same rate of evolution we see in the 50 million years that elapsed between Eocene and modern whales.
(N. Eldredge (1991) Fossils: The Evolution and Extinction of Species, Aurum Press, p. 168)
This is a problem: 100 million years before the Eocene takes us back before the evolution of mammals as a group (let alone the first mammals to take to the water).
Question 5
Can you identify a possible solution to this problem? (Hint: think about the assumption noted in the quotation.)
Answer
Perhaps the rate of evolution was not the same for the transition from terrestrial ancestor to primitive Eocene whale as it was for the later transition from Eocene whale to modern whales.
Simpson suggested that the rate of evolution had not been the same - that the large-scale transformations needed for a mammal to move from the land to water might have occurred more quickly than the subsequent 'fine-tuning' from primitive whale to modern whale. Some biologists see this as a general pattern, in which periods of rapid evolutionary change in small, isolated populations give rise to major new groups of animals, and are followed by longer periods of relative stability in which the rate of change to the new design is fairly modest. Niles Eldredge and the late Stephen Jay Gould have coined the term 'punctuated equilibria' for this controversial theory.
4.2 Intermediate forms
In essence, the argument about intermediate forms runs as follows. If whales evolved from a terrestrial ancestor through the accumulation of small differences over time, we should expect to find the fossils of a number of 'missing links', i.e. creatures with a mixture of terrestrial and aquatic characteristics. In fact, we might expect to find a succession of such animals, each a little bit more whale-like and a little bit less well adapted to life on land than its predecessor.
To make things more complicated, each of these intermediate forms must have been a fully working animal: it must have been able to breathe, to move about its environment, to feed itself and to reproduce. For a long time, biologists speculated about what these animals might have looked like. Surely at some point there must have been a creature that was at home on neither land nor sea, and so unable to compete with the animals already fully adapted to either habitat.
The problem of intermediate forms is by no means a new one. In The Origin of Species, published in 1859, Charles Darwin included a whole chapter on some of the difficulties facing his new theory. He started with the problem of intermediate forms:
Firstly, why, if species have descended from other species by insensibly fine gradations, do we not everywhere see innumerable transitional forms?
And a little later in the same chapter:
It has been asked by the opponents of such views as I hold, how, for instance, a land carnivorous animal could have been converted into one with aquatic habits; for how could the animal in its transitional state have subsisted?
Let's examine Darwin's questions one at a time. First, the scarcity of intermediate forms in the fossil record.
Question 6
What does the theory of 'punctuated equilibria' suggest about the chances of finding intermediate forms in the fossil record?
Answer
The theory suggests that we are more likely to find fossils of the animals that occupy the relatively stable periods (the 'equilibria') than we are to find the fossils of the animals that form part of the periods of rapid change, especially if that rapid change took place in small or isolated populations.
Now let's think about the nature of those animals.
Question 7
Can you think of any modern animals that are at home on land and in the water?
Answer
The TV programme contains a number of examples. Otters are wonderful swimmers and can chase and catch fish, but they also move about well enough to hunt on land [p. 186]. Seals and sea-lions are clearly most at home in the water, but they can still chase each other about on the breeding beaches. So it is possible to imagine an early ancestor of the whales leading a similarly amphibious existence.
Studies of the chemical make-up of proteins (particularly antibodies) in present-day species have shown that modern whales are closely related to the artiodactyls, a group of hoofed mammals that includes cows, sheep, deer and hippos. For some 100 years after the publication of The Origin of Species, all the known fossils of early cetaceans were either clearly terrestrial animals or primitive whales like those examined by Simpson. But recent finds have included the remains of creatures that might have had a more amphibious existence. These animals also lived in the Eocene (55 to 34 million years ago), in or around the warm, shallow waters of the Tethys Sea, an ancient ocean that stretched from modern Spain to Indonesia.
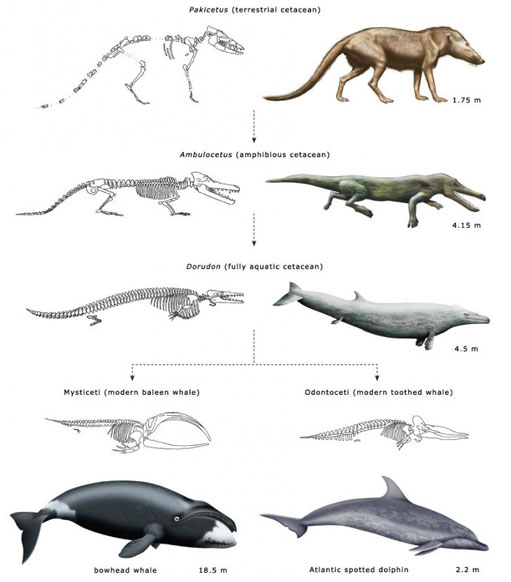
Figure 5 shows three representative animals in a postulated evolutionary sequence from an Eocene terrestrial ancestor to modern odontocetes and mysticetes.
Pakicetus is approximately 50 million years old. It lived by the side of shallow marine estuaries and was probably a wolf-sized ambush predator. It had long legs, an upright stance and may have spent much of its time running down small fish in the shallows.
Ambulocetus ('walking whale') is approximately 48 million years old, somewhere between Pakicetus and Dorudon. It was an amphibious animal with many of the characteristics we would expect in a transitional form. It could probably clamber about on land like a crocodile, but it was also a powerful swimmer, using its large feet and flexible spine like an otter. Intriguingly, it seems to have had a fluid-filled cavity linking its jaw and inner ear, similar to that found in modern dolphins (Figure 3), and it may have listened for the vibrations of approaching prey by resting its jaw on the ground - a strategy seen in today's crocodiles.
Dorudon is somewhere between 40 and 36 million years old. This animal lived in warm, shallow seas, where it fed on small fish and molluscs. It was about 4.5 m long and, like modern cetaceans, fully committed to a life in the water.
The Eocene was followed by the Oligocene (34 to 24 million years ago). The Tethys Sea disappeared as India ploughed into Asia, and the early whales found themselves having to live in deeper, colder waters. It didn't take them long, in evolutionary terms, to split into the two main lifestyles we see today. The toothed whales came to hunt fish and squid using high-frequency echolocation, while the baleen whales began to feed by filtering water, grew ever larger and sang their low-frequency songs across the oceans of the world.
Conclusion
This free course provided an introduction to studying Environment & Development. It took you through a series of exercises designed to develop your approach to study and learning at a distance, and helped to improve your confidence as an independent learner.
Acknowledgements
Except for third party materials and otherwise stated (see terms and conditions), this content is made available under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 Licence.
Course image: Ken Schneider in Flickr made available under Creative Commons Attribution-NonCommercial-ShareAlike 2.0 Licence.
Grateful acknowledgement is made to the following sources for permission to reproduce material in this course:
Reading 1: Phillips, H. (2001) Into the abyss, New Scientist, 31 March 2001, The New Scientist;
Reading 2: Reprinted with permission from Davis, R. W. (1999) Science, vol. 283, copyright © 1999 American Association for the Advancement of Science.
Figure 1 & 3 Bonner, W. N. (1980) Whales, Blandford Press Ltd;
Figure 2a Adapted from Bonner, W. N. (1980) Whales, Blandford Press Ltd;
Figure 4 Reprinted with permission from Davis, R. W. (1999) Science, vol. 283, copyright © 1999 American Association for the Advancement of Science.
Don't miss out:
If reading this text has inspired you to learn more, you may be interested in joining the millions of people who discover our free learning resources and qualifications by visiting The Open University - www.open.edu/ openlearn/ free-courses