Studying mammals: Life in the trees
Use 'Print preview' to check the number of pages and printer settings.
Print functionality varies between browsers.
Printable page generated Tuesday, 9 June 2026, 12:21 PM
Studying mammals: Life in the trees
Introduction
This course will introduce you to the wide-ranging types of mammals that live in the trees. You will learn how they thrive in this demanding environment, with the help of a range of intriguing adaptations related to their unusual life-styles.
This is the eighth in a series of units about studying mammals. To get the most from these units, you will need access to a copy of The Life of Mammals (2002) by David Attenborough, BBC Books (ISBN 0563534230), and The Life of Mammals (2002) on DVD, which contains the associated series of ten BBC TV programmes. This course contains samples from the DVD set. You should begin each course by watching the relevant TV programme on the DVD and reading the corresponding chapter in The Life of Mammals. You will be asked to rewatch specific sequences from the programme as you work through the course.
This OpenLearn course provides a sample of level 1 study in Environment & Development
Learning outcomes
After studying this course, you should be able to:
identify the common features shown by tree-dwelling mammals from different groups
show an awareness of the difficulties of classifying primates, especially in relation to the position of the prosimians
give an account of opportunities and challenges encountered by tree-dwelling mammals and of evolved adaptations linked with arboreal life
provide examples of the closeness (and sometimes interdependence) of the relationship between tree dwellers and their habitat
show an awareness of differences in lifestyle and forms of locomotion amongst a wide range of prosimians.
1 Life in the trees
As you work through this course you will come across boxes, like this one, which give you advice about the study skills that you will be developing as you progress through the course. To avoid breaking up the flow of the text, they will usually appear at the start or end of the sections.
As well as the course text, you will be using The Life of Mammals book (LoM) and related The Life of Mammals DVDs, as described in the introduction to this course. Before you go any further, watch 'Life in the Trees' on the DVD and read LoM Chapter 8. Unless stated otherwise, all the page references you encounter in this course will be to LoM.
The title of this course spells out exactly our focus of study; we'll be concerned with mammals that are arboreal - which means that to greater or lesser extents, they live in trees.
You won't be surprised to learn that tree-dwelling mammals don't comprise a single taxonomic group. A few familiar examples illustrate the point.
The striped possum is a marsupial. It's an agile, grub-eating tree dweller that uses its incisors to tear at wood and gnaw out wood-boring grubs. It has a sharp claw on an elongated finger, which can act as a probe or a skewer; it also has a long tongue, for probing the bark.
The binturong is the 'yard-long black hearth rug' referred to in Life of Mammals (LoM) p. 215. It's a civet - and therefore a member of the order Carnivora. It's an accomplished climber with a prehensile tail. Living mostly on fruit, it can hang upside down from branches - a lifestyle that is a significant departure from that of the more true-to-type carnivores.
You'll be familiar with squirrels as sure-footed tree dwellers - their agility is described and beautifully illustrated in LoM pp. 218-222. Squirrels are rodents - the description of squirrels as 'rats with bushy tails' has an element of biological truth.
The hyrax shows an adeptness at clambering around in trees. The middle area of each foot of the hyrax has muscles that can be retracted, so creating a suction pad that (along with rubbery soles, rich in sweat glands) help provide extra grip. Taxonomically, hyraxes belong to a group called the subungulates, which includes elephants and the aardvark.
What these examples reveal is that different evolutionary lineages have structures and lifestyles well-suited to the opportunities and demands of living in trees. In the remainder of this course, you'll come across numerous adaptations associated with tree dwelling. Many such examples - tails, claws, clinging feet - crop up again and again, revealing convergent evolution at work. Their similarity reflects the fact that the selection pressures that operate in such an environment are likely to be pretty well standard. But of course we'd expect a great deal of diversity too, in that very different types of mammals initially took to the trees. What's more, different species spend varying amounts of time in trees; some spend a good deal of time on the ground, others are on the ground fleetingly, simply to get from one tree to the other. So you'd expect tree dwellers to show varying degrees of adaptation to living in trees.
Activity 1
Watch the video sequence below showing climbing in the Malayan sun-bear and the tamandua - the South American anteater. (If you do not have time to watch the whole video, you should watch the section between 07:50 and 10:53 to see the Malayan sun-bear and the tamandua.) Think about the extent to which these tree dwellers are different from their terrestrial relatives. In each case, write two or three sentences that compare (a) the Malayan sun-bear with other bears and (b) the tamandua with other anteaters. Use the LoM index to find helpful information.
Answer
Distinctive features in these tree dwellers are not numerous! (a) The Malayan sun-bear has the strong forearms (and claws) of bears in general. But sun-bears are comparatively small, as you might expect of a climber. (b) The tamandua uses the powerful front legs typical of anteaters to climb, but its prehensile tail is distinctive, seemingly better suited for gripping by being more 'thinly furred' [p. 217]. But the arboreal pygmy anteater [pp. 60 and 217] also has a prehensile tail and compared to the tamandua this is more thickly furred.
2 Problems of life in the trees
2.1 The consequences of living in the trees
This course contains a lot of detailed information about particular tree-dwelling mammals. You will need to take care not to get too absorbed in the fine details but to ensure that you take away the important overall messages. Those are, of course, listed as the learning outcomes at the beginning of the course. Look now at the first one of those and bear it in mind while you are reading Section 2. You might find it useful to highlight or to make a list of the 'common features' as you come across them. Adopt the same technique with the remaining learning outcomes as you work through the rest of the course.
In this section, I'll be looking more deeply at some of the implications and consequences for mammals of living in trees, which will involve looking at the lifestyles of a range of examples. A good deal of what I say is descriptive 'natural history', much in the style that David Attenborough (DA) adopts in LoM, relating the lifestyles of tree dwellers to the environment in which they live. What we know of these animals reflects what is possible to observe in field studies, rather than from carrying out extensive investigation in the laboratory. Tree dwellers are elusive and cryptic animals, qualities that reveal how well adapted most of them are to tree dwelling and indicate how difficult the process of studying them under laboratory conditions has proved.
Question 1
Think about the 'hungry inquisitive animal' that DA opens Chapter 8 with [p. 213]. What problems are likely to be faced over time by such a mammal beginning to adopt a full-time tree-dwelling existence?
Answer
Moving effectively around trees requires agility and a good sense of balance. Dense foliage itself could also create problems of communication between individuals; reliance on vision may not prove an entirely workable strategy. Potential predators also pose a threat. Processing the most widely available food (leaves and fruit) may be far from straightforward.
How have tree dwellers dealt with such problems? Looking at a few examples of arboreal species in greater detail will provide some clues.
2.2 The kinkajou
LoM describes this tree dweller as a relative of the raccoon. It belongs to the order Carnivora and is one member of a family generally referred to as procyonids [p. 170], or more commonly the raccoon family. You'll be aware that some members of this family - for example, coatis [p. 174] - are omnivores. As you'll see in the video sequence below, coatis are more typically found in the undergrowth and leaf litter, rather than high up in the trees. (If you need to remind yourself of the lifestyle of coatis, go back to the previous video sequence and watch the section between 02:20 and 07:05 to see the coatis which feature at the beginning and end of the sequence.)
The kinkajou has a more restricted diet than coatis and is more committed to tree dwelling. It lives in the rainforests of Mexico and Central/South America. Superficially, it has some of the characteristics of primates - features that I'll say much more about in Section 3. For example, it has forward-facing eyes. As the photograph on p. 176 reveals, it also has a strong, monkey-like prehensile tail and clinging, clawed feet. These features allow it to hang down from branches, using dextrous forelimbs to reach the flowers and fruit that (along with a modicum of insects and small vertebrates) comprise its diet. So, in some ways kinkajous are monkey-like in their lifestyle, and yet they only half deserve the name by which they are known by local people - 'monkey of the night'. Indeed, they are nocturnal mammals.
Question 2
Does life in the trees guarantee protection from predators?
Answer
Certainly not, only from ground-based predators! The kinkajou is able to avoid the forest floor at night-time; this location is especially dangerous if jaguars are around [p. 146]. But tree dwellers are increasingly vulnerable to another set of predators, birds of prey. In daylight, kinkajous would be at risk from eagles.
The kinkajou's nocturnal habit helps ensure that predation rates are low. Forest owls might be a threat at night, but the kinkajou - at least the adults - are too large to fall victim to such a predator.
Animals like large herbivores that are heavily predated often live in herds where their strength in numbers mean that combined efforts can help to spot or perhaps fend off predators. For the kinkajou, predation is less of a problem, and may well explain why these animals are largely solitary in their feeding. Careful observations reveal that in more than 80 per cent of cases, individuals feed alone, which reduces the degree of competition between conspecifics, and is especially prevalent when food is less plentiful. Yet kinkajous do come together in well established social groups, just as primates do. In doing so, they display types of social behaviour, such as grooming and play between adults and juveniles, that are important 'bonding' interactions of primates.
Kinkajous have distinctive types of scent glands, located on their chin, throat and chest. Our understanding of what role these secretions play in communication in kinkajou society is very fragmentary, but they're seen as important in marking territorial boundaries between groups.
Question 3
Would it be appropriate to identify the use of scent as a straightforward adaptation to arboreal life?
Answer
To do so, you'd need evidence that this type of communication by smell was uniquely associated with tree dwelling, but this isn't the case. For many ground-dwelling carnivores, for example, smell plays a key role in social interactions; in the civet for instance [p. 142], territory marking by scent is very important.
But olfactory (as well as acoustic) communication often assumes a special importance in animal communities where visual contact is difficult, as would be the case for mammals living amongst dense foliage - and remember too that the kinkajou is nocturnal, so sight is not the most reliable of senses. But for those arboreal species that move swiftly - those that jump from tree to tree, for example - visual precision is needed, especially an ability to judge distances expertly. So the exact mix of the different senses shown by different tree-dwellers varies a good deal from one species to another, which is a factor worth watching out for as we look at other species.
2.3 The colugo
In LoM, DA vividly describes one particular evolutionary development associated with tree dwelling - taking to the air [pp. 221-227]. The gliding habit evolved independently in different mammalian lineages and yet the anatomical modifications that allow it are similar in, for example, flying squirrels and the unrelated colugo. In particular, the 'sail of skin' [p. 221], technically termed a patagium, stretches between the limbs - and a good deal further in the colugo, acting as an effective (and to some degree manoeuvrable) gliding membrane.
Colugos are sizeable mammals (as DA says, about the size of a domestic cat) and entirely arboreal. Their record-breaking glides (in excess of 70 m) are achieved without great loss of height. But in trees, they move about rather awkwardly. The patagium is then an encumbrance and there's a limited ability to grasp effectively - the colugo lacks the opposable thumb of primates that I'll talk about more in Section 3. So the benefits of a gliding lifestyle are achieved at a 'cost'. The resulting vulnerability - especially to the Philippine monkey-eating eagle (a species under threat, as are colugos) - may help explain why the colugo is nocturnal.
Taxonomically, the colugo is a bit of a mystery and the historical confusion is evident from its common name - the flying lemur. It neither flies (in that it doesn't flap its limbs) nor is it a lemur (see Section 4). Neither, of course, is it a monkey, despite the name ascribed to its main predator. Having once been placed in the insectivores and then in the bats, it's now ascribed a mammalian order of its own (the Dermoptera, i.e. 'skinwings'), recognising its ancient and distinct evolutionary beginnings. You'll appreciate from LoM p. 223 why few clues as to the colugo's evolutionary affiliations emerge from a study of its remarkable dentition. Its specialised stomach is able to deal with large amounts of leafy vegetation and the large intestine is disproportionately large.
Activity 2
Flying squirrels are not closely related to the colugos but they have features in common. Watch the video sequence below. (a) Identify one similarity and one difference between flying squirrels and colugos. (b) On the evidence of the video sequence, comment briefly on how flying squirrels steer during gliding. (c) Write a sentence or two about the disadvantages and consequences of the gliding habit in flying squirrels.
Answer
(a) The patagium is the most obvious similarity, though it is less extensive in flying squirrels - compare the photographs in LoM pp. 221 and 222. The two mammals differ in that the tail in flying squirrels is free of the patagium and said to act as a rudder in flight, whereas in the colugo, the tail lies within the patagium.
(b) The presumed steering ability of the tail is difficult to discern. Manoeuvrability in flight seems more a consequence of adjustments in how the patagium is positioned. (Braking during landing is said to be achieved by upwards movement of the tail and body, though I couldn't see this on the few examples of landing shown in the video sequence.)
(c) The possession of a patagium limits climbing in the trees (though these flying squirrels seem to scamper about unhindered). Their nocturnal habit is probably related to increased vulnerability during gliding.
2.4 Fruit bats and flying foxes
In this section and the next, you will be asked to write answers to activities in particular numbers of words - Activity 3 asks for about 200 words and Activity 5 requires 150 words - you might like to use your Learning Journal for this. Guidance of this type gives a useful indication of the degree of detail required in your answer. It is important that you continue to attempt these activities yourself, without looking ahead at the answer and that you then critically compare your answers with mine. It is quite possible to write the specified number of words but not to include all the essential points. So, a useful exercise is to identify each point in my answer and check whether you have included it in yours. If you have missed out any points, is that because you made a conscious decision to exclude them, or because you were just not aware of them from your reading? If the former, then consider whether you could have written the remaining points more concisely to allow everything to be included. If the latter, then you may need to adapt your study technique to do more highlighting, or make more notes, so that you are certain that you have identified all of the relevant points.
Colonies of flying foxes may comprise as many as a few million individuals (five million is DA's estimate), each with a wingspan of about 1.4 m, with the entire 'camp' perched on often denuded trees and engaged in intense social activity.
It's little wonder that witnessing such a site has been described as a 'memorable auditory and olfactory experience'.
Activity 3
Watch the video sequence below on flying foxes. You might find it useful to reread LoM pp. 223-227. Discuss, in about 200 words, the likely advantages to flying foxes of their particular form of roosting, taking into account vulnerability to predators, the location of food and temperature regulation.
Answer
Such concentrations of flying foxes are 'visible, audible and smellable for miles' [p. 227] and therefore inevitably attract predators. But congregations of this type may decrease the likelihood of any one individual falling prey to predators, such as eagles. Communication between members of the camp may also increase the efficiency of locating suitable food. But the fact that food sources are depleted so comprehensively when visited en masse raises questions as to the degree of benefit of group living to any one individual fruit bat. Another possible benefit of roosting is that foliage might be protective, shading these mammals from wind, rain and sun, though trees that become camps lose many of their leaves. Bats regulate their body temperature, partly by behavioural means. Huddling together in groups should in theory reduce the rate of heat loss in cooler conditions, and decrease the rate of warming when it's very hot. In both circumstances, the surface area that each individual exposes is lessened by contact [see p. 224], though this idea implies a degree of restful intimacy not entirely consistent with DA's descriptions and the video evidence of continual jostling.
As the video sequence showed, eagles (and owls) take a toll of flying foxes in transit, and the largely nocturnal habit of these species once again probably reflects selection pressure of this type. Flying foxes living on islands (more than 60 per cent of species do so) tend to venture forth in the daylight and in such environments predators are often less evident.
LoM highlights the devastation that flying foxes can bring to a farmer's crop [p. 227]. But DA also stresses that they can maintain 'the fertility of the rain forest'. Flying foxes can certainly help disperse trees by transporting their seeds to new locations, either through their messy eating of fruits or by seeds passing intact through the gut. The seeds of the commercially important West African iroko tree depend on the straw-coloured flying fox for their dispersal. Flying foxes also help in the recolonisation of deforested areas and in the establishment of plants on land newly formed or recently devastated by volcanic eruption.
Flying foxes are also important pollinators; many island species occupy the ecological niches taken over elsewhere by insects or humming-birds, for example. The transfer of pollen from one flower to another on a different tree (i.e. cross-pollination) can confer a significant advantage to the species because it promotes genetic diversity of the next generation. So the development of mechanisms that promote cross-pollination are very advantageous to trees. In Australia, pollination of some eucalyptus species depends almost entirely on visits from flying foxes. The flowering process of the Kajeng Jaler tree from Malaysia is intimately geared to the feeding habits of the dawn bat. Its flowers open just two hours or so after dusk and drop before dawn, coincident with the bat's feeding time. The size and shape of the flower opening ensure that only the dawn bat can enter; as its long tongue reaches down to access the nectar, the position of the pollen-producing parts of the flower (the stamens) is such that pollen is deposited on the animal's fur.
This is a further demonstration of the way in which the evolution of one species can increase its dependence on another, often reflecting some form of mutual advantage. This is phenomenon is known as coevolution.
2.5 Tree squirrels
Coevolution also underpins the relationship between many tree squirrels and the trees that house them. The creation of food caches as a 'winter-larder' is mutually beneficial, partly because squirrels are sufficiently profligate in their habits to ensure that many stores are overlooked. Stealing by neighbours is so common that such over-provision may be essential - it's not through forgetfulness or lack of skill; grey squirrels appear able to detect nuts buried as deep as 30 cm below the surface. As you know, seeds that remain buried are often well placed to germinate. But neglecting some caches is a viable tactic only if an abundant crop of seeds is produced. The sacrifice of a modest number of seeds to satisfy the squirrel's habits is seemingly a cost worth paying.
But this cosy mutuality is not guaranteed. The tassel-eared squirrel, for example, depends on the Ponderosa pine for food and shelter. Oddly, this species has never acquired (or has lost over evolutionary time) the habit of cache-making. Neither does it undergo winter hibernation - though remaining active throughout the year is far from unusual amongst squirrels. In the winter months, this squirrel exists almost exclusively on a diet of the inner lining of the bark (the phloem layer), which is an essential component of the tree's transport system; the resulting damage to the tree is significant. The benefits from this tight relationship are seemingly very one-sided.
Tropical tree squirrels provide an excellent example of niche separation. Different species don't mill around indiscriminately in any one area of the tropical rainforest; rather, they have distinct lifestyles and preferences for particular foodstuffs. The background of tree outlines in (below) shows the various layers of tropical rainforest - for the moment, ignore the placement of the different species of tree squirrel. The trees comprising the canopy grow up to 40 m in height, their growth producing a continuous and dense mass of overlapping branches, leaves, flowers and fruit. Some taller trees (emergents) can burst through the canopy. In the middle layer, the trees are somewhat shorter and the vegetation less dense. The understory comprises small and young trees, while the forest floor is a dark area of shrubs and ferns, plus a layer of leaf litter, which is subject to rapid breakdown and utilisation via the action of decomposers and scavangers.
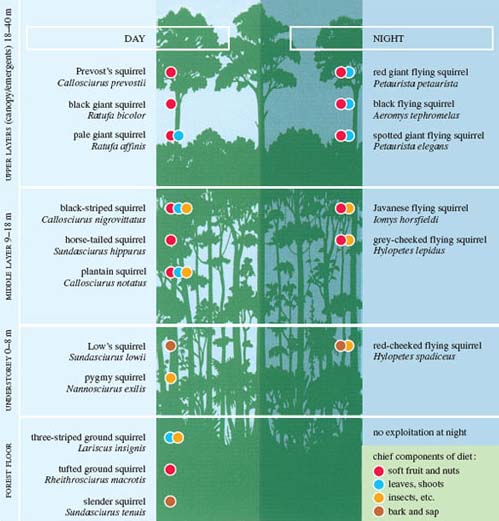
Question 5
Question: Which layer in Figure 1 is likely to be subject to very changeable physical conditions and to receive the greatest sunshine and heaviest rainfall?
Answer
The uppermost canopy layer, i.e. the crowns of the tallest trees, plus the growth that rises up through it - the emergent layer.
Question 6
Question: Look at the location of the middle layer. What conditions are likely to prevail here, with respect to temperature, air movement and rainfall?
Answer
This layer is more protected from sunshine and is generally more stable. For much the same reason, there is relatively little air movement. Rain from the canopy eventually drips downwards and the still air can readily become saturated with moisture.
In one particular Malaysian forest location, 25 species of tree squirrels have been identified, 11 nocturnal and 14 active by day (i.e. diurnal). (Relatively few Malaysian squirrels are seed eaters, compared to their African or temperate forest counterparts.)
Activity 4a
Study the information in Figure 1 showing where many of these tree squirrels feed and their food preference.
(a) Does any one species feed during both the day and the night?
Answer
No; as you'd expect these species are either diurnal or nocturnal, i.e. there is separation between these species in time. (Note that the flying squirrels are all nocturnal.)
Activity 4b
(b) Does any species feed in more than one part of the forest?
Answer
No, each species seems to be restricted to a particular forest layer. Flying squirrels tend to feed in particular locations - for example, the upper or middle layers - which means there is separation between these species in space.
Activity 4c
(c) Do the species that inhabit the same part of the forest profile generally eat different foodstuffs?
Answer
Sometimes diets are distinct and the species non-competitive - as in the three diurnal species feeding on the forest floor. But there is some overlap; for example, look at the three diurnal species feeding in the upper layers.
Activity 4d
(d) What conclusions can you draw from the information in Figure 1? Write an answer of about 50 words.
Answer
The forest provides a variety of resources available for exploitation - e.g. insects, fruit, leaves and bark - located in different parts of the profile. Differences in the lifestyles and feeding preferences of tree squirrels reflect their occupation of particular niches, and may reduce, though not eliminate, competition between species for these available resources.
This is an example of niche separation, which in this instance may be even more marked than Figure 1 alone suggests. The three diurnal squirrels identified in the upper layers have subtly different fruit preferences and feeding strategies. For example, Prevost's squirrel has a narrower range of preferred fruit, and its smaller size and greater agility enable it to reach foods inaccessible to the black giant squirrel. But niche separation works best when foodstuffs are relatively plentiful. With these Malaysian tree squirrels, nearly all species consume bark and sap when other foodstuffs are less freely available, at which point direct competition between species is more prevalent.
Activity 5
As a conclusion to what you have read in this course period so far, give a brief account, in about 150 words of the consequences of the tree-dwelling habit for mammals, with examples. Refer in your answer to the key concepts of the influence of predators, sensory communication, coevolution and niche separation.
Answer
Tree-dwelling mammals may escape terrestrial predators but they are exposed to others, notably birds of prey. Many tree dwellers are nocturnal and there are advantages for some (e.g. flying foxes) in congregating in large numbers, which may have the advantage of decreasing the risks of predation to any one individual. Since visibility in trees may be limited, communication by smell can be important (e.g. in the kinkajou), though judging distance requires visual acuity. A reciprocal influence on evolution is evident in the intimate relationship between some arboreal species and the trees that sustain them, e.g. pollination by flying foxes reveals coevolution. Tree dwellers are unlikely to feed indiscriminately; rather there is niche separation. Different tree squirrels that occupy the same part of the forest may have different food preferences. Available niches within a forest area are likely to be utilised by specialist feeders.
3 Introducing the primates
One group of accomplished tree dwellers are the primates - a term you perhaps think of as synonymous with monkeys and apes. Monkeys and some apes display some of the most striking adaptations to tree-living.
250 or so species of primate exist today; most taxonomists group them into 13 families. All share a lengthy list of defining features, mostly related to the following broad categories:
Limbs and locomotion. The hands (and often the feet) are grasping, with mobile fingers and toes, generally with touch-sensitive pads at the tips. The first digit (the thumb or the hallux, i.e. big toe) is normally divergent (i.e. points outwards) and in many species can be swivelled to bring its tip into contact with other digits; in other words, it is opposable, to a greater or lesser degree. (Try this with your own hand to verify it.) Rather than a curved and rigid claw, at least some of the digits of primates have flat nails, making manipulation (e.g. of food) practicable. They have very flexible shoulder joints; hindlimbs are normally dominant in locomotion.
The senses. The eyes point forwards and are set close together. The fields of view of the eyes overlap which, together with a distinctive 'wiring' arrangement linking the eyes and the brain, imparts stereoscopic, '3-D' vision. In general, vision (like hearing) is more significant than smell, and colour vision is widespread. The face is flattened, with the muzzle (i.e. the area of skin around the nostrils) foreshortened; this anatomical change may, in part, be a reflection of the diminished importance of smell.
The brain. This organ is relatively large. The cerebral cortex - in primates, more often called the neocortex - is elaborately folded and complex in structure. This part of the brain is involved with the highly complex processes that include learning, reasoning and memory.
In all, up to 30 or so diagnostic features of the type described above have been identified for primates, though not all primates display each trait. The others include: their distinctive dentition, linked with a generally omnivorous diet; the structure of the ear; and reproductive features, such as small litter sizes and long gestation periods, relative to body size. Primates have an extended period of juvenile growth and their overall rates of growth and reproduction are generally low, which some biologists interpret as the price paid for a large brain. Brain tissue is metabolically a very expensive tissue to develop and maintain, so for large-brained mammals, less energy is available for growth and reproduction. The benefits of a large brain are very significant. For example, the elaborate social behaviour of primates is seen by many biologists to be as much a defining feature of primates as the types of anatomical feature just listed.
While monkeys and apes (and ourselves) may be the most familiar primates, they are far from the only members of this order. I'll focus on types of primates other than apes or monkeys, where again a tree-dwelling lifestyle is especially widespread. Such primates comprise three subgroups, all introduced in LoM:
the bush-babies (or galagos) of Africa (Section 4);
the lorises of Asia and the pottos of Africa (Section 4);
the lemurs of Madagascar (Section 5).
LoM refers to all such primates as prosimians [p. 230]; by contrast, monkeys and apes are termed 'anthropoids'. The term prosimian implies that these primates are in some sense 'forerunners of the monkeys' - historically monkeys and apes were referred to as 'simians'. More than likely, prosimians do more closely resemble the ancestral primates than do modern-day monkeys and apes, though thinking of them simply as primitive 'pre-monkeys' would be a mistake. As you'll see, they have complex and rich lifestyles and display sophisticated adaptations to tree dwelling.
LoM mentions one type of prosimian that stands apart from its relatives [pp. 243-245], the wholly carnivorous tarsiers, and attention is drawn to their rather anomalous taxonomic status. Indeed, most taxonomists now favour grouping tarsiers within the primate suborder that comprises the true monkeys and apes and urge the abandonment of the term prosimian. You're likely to encounter this preferred classification scheme in your wider reading and in the classification scheme featured in the Open University course from which this course originates (S182), so I'll outline it here.
The newly defined suborder Strepsirhini includes what DA refers to as prosimians, i.e. galagos, pottos, lorises and lemurs - but excludes the tarsiers. (The suborder takes its name from the features of the nose, in particular a naked area (the rhinarium) surrounding what are often referred to as 'comma-shaped' nostrils.) The muzzle of strepsirhines is commonly described as 'prominent, moist and glandular' and, as I've hinted, strepsirhines are thought, for a variety of reasons, to resemble more closely the ancestors of primates. Compared to other primates, they have smaller brains, a better sense of smell and longer snouts. As DA's comments imply, the tarsier's muzzle is different: 'fur grows almost to the edge of them [the nostrils] and surrounds them' [p. 245], i.e. the muzzle is hairy. This feature defines the suborder Haplorhini, together with the fact that the muzzle is less prominent. The haplorhines therefore include the tarsiers and the monkeys and apes.
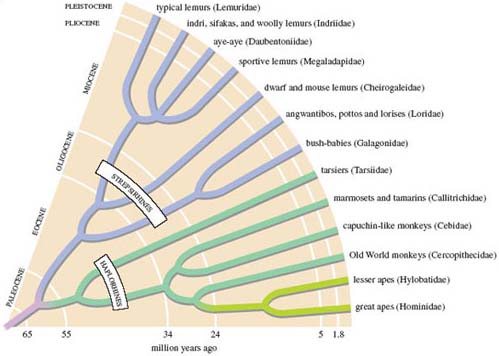
This revised classification is meant more accurately to reflect the presumed evolutionary history of primates. Figure 2 provides such an evolutionary tree, plotting the different lineages that evolved from the presumed common ancestor of primates, stretching back into the Paleocene. You can see a divergence from this ancestor into the two branches reflecting the suborders just introduced, the differences in colour evident as they split. Continued branching of each of these evolutionary lines, at different periods in the geological past produced - perhaps as much as 100 million years or so on from the ancestral primate - the range of 13 families that comprise today's primates. But for consistency with LoM, I'll refer to the two suborders of primates as prosimians and anthropoids, rather than using the haplorhines and strepsirhines division of Figure 2.
I've already drawn attention to the distinctive hands and feet of primates. shows some of this anatomical diversity in primates, including representatives of the monkeys and apes. The hand of a tree-shrew, i.e. a non-primate, is shown for contrast (Figure 3a); this climbing mammal is not particularly well adapted for living in trees (some species of tree-shrew are almost entirely ground-dwelling). Tree-shrews have claws on all their fingers and their thumb - and the thumb is hardly different from the other digits. The digits are able to do little more than spread out or converge, rather than span and grip. In contrast, the feet and hands of the primates shown in Figures 3b-j generally have nails, which are straighter and less robust than claws. They also have tactile pads and a more manoeuvrable thumb and toe - features that are used to good effect in the tree-climbing species illustrated here. Figure 3 reveals the great anatomical diversity of primate hands and feet and a widespread ability to grasp, i.e. they are very often prehensile, which is reason enough for us to come back to the figure later in this course.
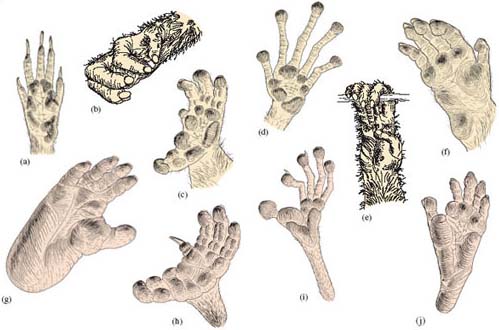
In Figure 3 (a) is the paw of a tree-shrew, for contrast. (b) shows the precision grip of the chimpanzee, reflecting their higher degree of manual dexterity. (c) and (h) are the hand and foot, respectively, of a potto; (d) and (i) are the hand and foot, respectively, of a tarsier. (e) is the hand of a gibbon; the thumb is short, highly opposable and distant from the remaining digits, which are slender and well suited to grasping. The arm-swinging form of locomotion of gibbons, which is called brachiation. The relatively long forelimbs, together with anatomical adaptations that increase the mobility of the shoulder, forearm and wrist, contribute to this very efficient form of locomotion, ideally suited to an environment where handholds are irregular and widespread. (f) is the hand of a macaque monkey, which is adapted for walking with the palm flat on the ground. (g) and (j) show the foot of a gorilla and baboon, respectively, both largely ground-dwelling primates.
4 Galagos, lorises and pottos
Activity 6 is divided into several parts, on successive pages, and asks you to answer a number of questions. Look ahead to this activity now and highlight, or list, the different 'process' words, which tell you what you need to do with the content You should have found 'describe', 'contrast', 'evaluate' and 'explain'. If you look back over earlier activities in this study period, you will also find 'compare', 'discuss' and 'give a brief account of'. You may find it useful to look up such terms in a dictionary, which would help you to work out what is required of the answer. For example, 'describe' means give an account, 'contrast' means compare in order to show differences, and to 'evaluate' requires making an appraisal of something's worth. If you were asked to 'discuss' something, that means you should investigate or examine by argument, sift and debate, give reasons for and against.
Galagos, lorises and pottos were introduced in Section 4; Figure 2 suggests that they derived from a common ancestor, which diverged about 20 million years ago and now forms two families. Pottos and lorises are more closely related to each other than to galagos. Nearly all are tree dwellers, as their hands and feet reveal. Generally, the thumb and often the big toe are enlarged, diverge strongly and are to some degree opposable on the hand or foot. This feature, together with a second digit that is reduced to a small stub, ensures a wide span and an ability to grip branches. The hands and feet of the potto in Figure 4c and h show all these features; those of the loris, which DA describes in LoM p. 228, are much the same. An unusual dentition helps define the group: in particular, lower incisors and modest canines project forward, forming a toothcomb structure, which is involved in both grooming and in some specialised forms of feeding. In fact, nearly all prosimians have a toothcomb of sorts - the main exception being the tarsiers - and in some species (e.g. Figure 7c) it is especially prominent. Its action normally combines with that of a horny plate under the tongue (the sub-lingua), which helps remove debris such as hair that accumulates within the toothcomb during grooming. Another diagnostic feature of the group as a whole is the 'grooming' claw on the second digit of each foot (Figure 4h).
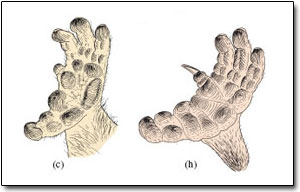
Galagos (bush-babies) are restricted to Africa and the lorises inhabit India and Southeast Asia. (I'll say very little about the pottos; DA refers to them briefly in LoM p. 230. I'll also ignore the angwantibos of West Africa, although they are in Figure 2.) Activity 6 highlights a few key points about these prosimians, via a number of questions. After several of the answers, I'll expand on some of the points made.
Activity 6a
To answer the following questions, you may find it useful to study LoM pp. 227-234 again, including the illustrations, and to watch the video sequence below.
(a) In one sentence, describe the diet of lorises and bush-babies.
Answer
Both lorises and bush-babies are primarily insect eaters, though both supplement their diet, often with fruit such as berries.
Some bush-babies are specialised for feeding on the gum that flows from trees that have been damaged - by insects, for example. The toothcomb allows most gum-feeding bush-babies simply to scrape off exuded gum, to supplement other food sources. But the needle-clawed bush-baby is more dependent on this food source. It has modified canines and premolars that allow it to gouge holes in the bark; this strengthened toothcomb is shown in Figure 7c. The gum of trees contains complex sugars, which the usual repertoire of digestive enzymes is relatively ineffective at breaking down. So these bush-babies have a form of microbial fermentation similar to that in some herbivores. The microbes that carry out the fermentation are housed within the caecum (an offshoot of the large intestine), which in the needle-clawed bush-baby is five times bigger than expected for a similar sized mammal with a more conventional diet. (Feeding on gum is a widespread habit amongst tree dwellers.)
Activity 6b
(b) In three or four sentences, contrast the methods of locomotion of lorises and bush-babies.
Answer
Lorises have a creeping form of locomotion - almost a type of unhurried stretching, via limbs of approximately the same size. Their hands have a very wide span and they are able to keep a forceps-like grip on branches. Bush-babies also have grasping hands, but they are specialised leapers and their hindlimbs are considerably longer than their forelimbs [see the photo on pp. 232-233] and their bushy tails are said to act as stabilisers during their leaps.
Given their permanent location on branches (often high up, in the canopy) the forceps-like grip of lorises is a vital adaptation. It requires sustained muscular contraction and the 'special mesh of blood vessels in its wrists and ankles' that LoM p. 228 refers to underpins that ability; it's more formally termed a rete mirabile (pronounced 'rett, or 'retay', mirabilay', and Latin for 'wonderful net'), or simply rete. DA links this rete to an ability to maintain a grip. How might this happen? Highly active muscles require a continuous supply of oxygen and blood-borne nutrients; muscle fatigue can be minimised by the removal of lactic acid. The accumulation of lactic acid limits the action of our own muscles during heavy exertion; it is also associated with deep prolonged dives in aquatic mammals. In all such events, it is a consequence of anaerobic metabolism, brought into play when a reduced oxygen supply is limiting the ability of muscles to contract. The local accumulation of lactic acid would alter the acidity around the contracting muscles and render them ineffective; the rete mirabile perhaps helps transport the lactic acid away, to be dealt with by the liver. Most examples of a rete that you're likely to come across promote the exchange of heat, but the rete here is an example of unusual biological engineering!
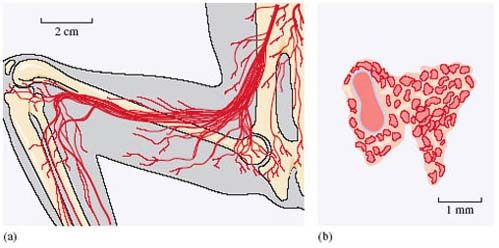
Figure 5a (below) shows the rete mirabile in the thigh region of a slow loris. What's evident is a mesh of finely branched, narrow blood vessels - a more conventional arrangement would be very much fewer and larger vessels, where a few major arteries would convey blood towards the feet and a few veins convey blood back towards the core of the body. The intricacies of the rete mirabile are revealed in the cross-section in Figure 5b, which is of the comparable structure in the potto. A major artery (the brachial artery) is surrounded by a lattice-work of smaller arteries and veins. The anatomical arrangement is very reminiscent of structures primarily invlolved in the countercurrent exchange of heat, but a different mechanism seems to be at work here. An array of smaller arterial branches of this type may give the entire bundle of vessels a degree of rigidity. The veins containing the outflowing blood are less likely to collapse during sustained muscular contraction, as they might do in more conventional arrangements. Keeping the blood flowing in such circumstances avoids the accumulation of lactic acid, enabling the grip to be sustained.
Activity 6c
(c) Evaluate the following statement: 'Urine washing in lorises and bush-babies is known to increase the grip of the hands and feet.'
Answer
The function of urine washing is uncertain and this statement doesn't recognise alternative or additional explanations. Urine washing may be linked with territorial marking, much as may be the case in the kinkajou. Some researchers claim that urine washing has a cooling function; others suggest that urine has a disinfectant role. Further investigation would be needed before the function of urine washing in lorises could be stated with such certainty.
Activity 6d
(d) Some experts have argued that stereoscopic vision of the sort possessed by bush-babies is essential for estimating distances in arboreal leaping. But what tree-dwelling mammals encountered earlier in this study period have laterally-pointing eyes and yet are seemingly expert leapers?
Answer
Squirrels provide perhaps the best example [p. 219]. Their laterally-positioned eyes ensure a broad field of view, but they appear not to have a significant degree of stereoscopic vision. It may be that bush-baby-style vision is important for hunters - think of owls, hawks and cats - which squirrels are not.
Activity 6e
(e) Explain the function of the tapetum lucidum in the eyes of lorises and bush-babies.
Answer
For nocturnal species, the tapetum lucidum maximises the sensitivity of the eye to low light conditions, by reflecting back into the retina light that has initially passed through, providing a 'second-chance' for activation of the light-sensitive receptor cells.
There are two major types of light-sensitive cell located within the retina of mammals, different in shape, chemical composition and function. Rods enable species to see in black and white; the presence of cones provides colour vision. By working out the ratio of rods to cones in the retina, an animal's ability to detect colour can be established. As you'd predict of nocturnal animals, pottos, lorises and bush-babies appear not to have colour vision. In prosimians generally, the proportion of cones to rods within the retina is low.
5 Lemurs and their origin
5.1 Introduction
The island of Madagascar is relatively small compared to the enormous landmasses of Africa, Asia and South America inhabited by the primates I've mentioned up to now. Yet of the 250 or so living primate species, more than 30 species of lemur live in Madagascar - an astonishingly high 13 per cent or so of all primate species. Their diversity is reflected in the identification of no fewer than five families, related in the way suggested in Figure 2.
What are just as striking are the mammalian groups that are absent from Madagascar or sparsely represented - there are none of the large African herbivores, few carnivores, no rabbits or hares and no monkeys or apes; lemurs have evolved in an environment with a unique mix of fauna. All lemurs could loosely be described as tree dwellers, usually to a very marked degree, and display a correspondingly rich array of adaptations. As our knowledge increases, so does the species count of lemurs; species thought to be extinct have recently been rediscovered. Careful fieldwork has revealed subtly distinct species coexisting in what was previously thought to be a homogeneous population. Genetic evidence, e.g. numbers of chromosomes or DNA sequence comparisons, has also led to further subdivisions. If you add to the score the many lemur species that have become extinct over the past few thousand years, the total number of evolved species is close to 50.
As I've mentioned, lemurs are the only primates on Madagascar (humans apart) and their evolutionary development over the past 50 million years or so has taken place in an isolated, self-contained 'natural laboratory' of evolution. Genetic evidence has shown - more convincingly than morphological evidence is ever likely to - that lemurs are monophyletic, meaning that they are share a single common ancestor, which according to Figure 2, would have lived in the Eocene. What is striking is the subsequent 'bushiness' of the evolutionary tree, in the major part of the branch labelled strepsirhines. This proliferation of species reflects the process of adaptive radiation: over time, a single lineage has diversified into a number of evolutionary branches that occupy different niches and have a distinctive mix of adaptive features.
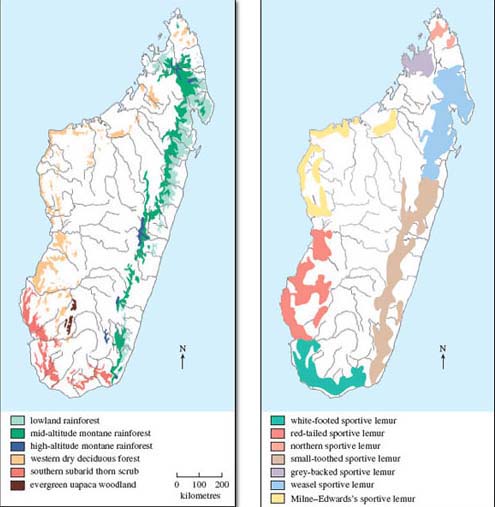
Quite why so many different species have evolved in a relatively small landmass isn't known for certain. It may be linked to the different climate zones on the island, each with distinctive plant species, and to the influence of many natural barriers in the form of rivers and land of very varying altitude - features that would tend to isolate populations and thereby promote the origin of new species (i.e. promote speciation). The diversity of environments Madagascar contains indeed rivals that of larger continents, though the areas of untouched vegetation are now a fraction of what they used to be, as Figure 6a implies. Much of the island nowadays is agricultural grassland. Of the original vegetation, representing the major habitats for today's lemurs, there are (montane, i.e. high altitude) rainforests in the east and deciduous forests in the north and west; in the southwestern parts of the island there is dense thorn scrubland, plus vestiges of evergreen woodland.
The enormous diversity of lemurs reflects the extraordinary scale and power of the mechanisms that drive evolution. For example, lemurs vary greatly in size; the mouse lemurs [p. 234] are tiny - indeed the pygmy mouse lemur - weighing just 30 g or so - is the smallest known primate, about the mass of a house mouse. Giant sloth lemurs are now extinct, but were in excess of 200 kg - larger than a modern-day gorilla. The variation in dental formulae amongst lemurs (and the morphology of the molar teeth) is said to be greater than in all other living primates taken together. And as you'd expect, there is considerable specialisation in lemur eating habits: some eat leaves; others, like the ring-tailed lemur [p. 240], prefer fruit. The diet of bamboo lemurs is self-evident; in fact, the three different species identified feed on different parts of the plant; for example, the golden bamboo lemur (which you will see in the next video clip) feeds just on new shoots, which contain amounts of cyanide that would be lethal to other primates. Other lemur species eat gum or small invertebrate prey. There are wide variations too in the social groupings that lemurs form, which are often different from the patterns evident in monkeys and apes. In general, group sizes in lemurs are smaller than in monkeys and apes and females tend to be the dominant sex.
Overall, lemurs are very distinctive and unusual primates. Primatologists assume that their odd features - their social groupings, less constant body temperatures, and low metabolic rate, for example, - are adaptive, i.e they increase fitness. Explaining why such adaptations are beneficial is problematic, when other primates thrive on opposite strategies. Most explanations link them with the rather frugal existence that lemurs endure on Madagascar - the climate, and therefore food supply, are unpredictable and the soils are poor. Lemur evolution seems to have been influenced very sharply by the need to maximise the use of scarce resources and the need to conserve energy.
Madagascar split away from mainland Africa in the distant past [p. 234]. The best estimates are that the process was complete by about 120 million years ago. It's likely that the first primates evolved more recently - perhaps as long as 90 million years ago, but certainly after detachment of the island. Clearly, no early lemur could have migrated overland on to what is now Madagascar; you can check from Figure 2 that lemurs originated at a much later date, when Madagascar was an island. And yet nearly all expert opinion claims that the ancestors of lemurs originated from Africa, rather than, say, what is now India, from which Madagascar finally severed its connection more recently - about 88 million years ago. So how could the ancestors of lemurs have arrived on an isolated Madagascar?
The most widely accepted explanation is that the first primates arrived by floating across from Africa on mats of drifting vegetation, perhaps 60 million or more years ago. The Mozambique Channel is more than 400 km wide, so the journey would have been a significant feat of endurance, but given the presumed monophyletic origins of lemurs, we can view it as a rare but fortuitous event.
Question 7
What attributes in these early primate 'sailors' would have increased their chances of success?
Answer
Being small and light would presumably be advantageous, helping them to stay afloat on small bits of driftwood, etc. Indeed, modern-day lemurs that most resemble ancestral forms are small - the dwarf and mouse lemurs [p. 234] - and are generally regarded as the 'best living models for ancestral primates'. You can see in Figure 2 that this group and the remaining lemur families parted evolutionary company more than 30 million years ago.
Question 8
What feature of small mammals (linked with their high metabolic rate) would be a severe disadvantage in 'rafting'?
Answer
Their high metabolic rate is reflected in their voracious appetites, prompting them, as with shrews, to spend a large proportion of their waking hours feeding.
As I've hinted already, many lemurs have a lower than expected basal metabolic rate (BMR) but torpor is prevalent too. For example, when climatic conditions are unfavourable, normally during the cool summer, dwarf and mouse lemurs can spend days in torpor - a feat that no other primate appears able to match. Indeed, days can sometimes stretch to weeks and months, with the animals reducing their metabolic rate by up to 90 per cent and lowering their body temperature close to ambient. Recent field studies have shown that these lemurs usually aggregate during torpor, in huddling groups of up to 15 in single tree-holes. Keeping each other warm in this way increases the energy saved during torpor yet further.
All these capacities have been used as evidence to support the rafting hypothesis in that it suggests a plausible mechanism for this feat of endurance. At one stage, the belief prevailed that colonisation involved 'a pop-eyed ancestral lemur clinging with all its limbs onto the wave-washed twigs of a raft of floating tree trunks or tangled branches'. Now the picture painted is of 'entire groups of animals survived the weeks or even months of such a journey without food or water, sleeping in a hollow tree while rafting across the sea'. Both are wonderfully evocative images, given the inheritance that these pioneers have left behind. Perhaps perspectives on this issue will change in the years ahead - possibly with renewed support for some of the alternative hypotheses for migration, such as island-hopping across the Channel after a temporary reduction in sea-level.
And what of human influences on lemurs? Here the tale is depressingly familiar. The first human settlers arrived in Madagascar about 2000 years ago - probably from the Malaysian/Indonesian area. By the time Europeans arrived in the country in the mid-1660s, 15 species of lemur had become extinct, amongst them the largest, most striking and bizarre, such as the sloth lemurs.
5.2 Madagascan diversity
Activity 7
Watch the video sequence below, which focuses on just three lemur species - the ring-tailed (in a very brief sequence, leaping from one tree to another), the golden bamboo lemur, already mentioned, and the sifaka, plus the fossa. (a) Write down in about three sentences your preliminary observations on the differences in lifestyle and means of locomotion between these lemurs. You may find it useful to reread LoM pp. 234-241. (b) How would you classify the fossa, in taxonomic terms and by feeding habit? Has anything like it been mentioned so far in this course?
Answer
(a) Ring-tailed lemurs are accomplished tree-dwellers, clearly able to leap from tree to tree. The golden bamboo lemur is distinguished by its remarkably specialised (and toxic) diet; it is clearly able to cling to the thick stems of bamboo. The sifaka's form of locomotion (along with that of the indri) is highly specialised - an athletic leaping - and a distinctive balletic style of movement on the ground.
(b) The fossa is a carnivore and is unique to Madagascar; it preys on lemurs - the clue was in the video commentary, where the mammal was described as a 'kind of giant mongoose'. It's a civet - a member of the family Viverridae. It therefore belongs in the same group as the binturong, mentioned in Section 1 and LoM p. 215, but is very different from it in lifestyle. The binturong is a frugivore and 'moves in a very leisurely way'. Looked at superficially, the fossa more obviously resembles a large cat - equally at home in trees and on the ground. Over many millions of years, the fossa's isolation from other carnivores and its life within this distinctive fauna has meant that this civet has followed a unique evolutionary track.
DA's description indicates the great range of behaviours and habits in lemurs; I want to add a few more points. Refer back to Figure 2 at intervals for more information on the affiliations of each of the lemurs mentioned.
5.3 Sportive lemurs
Sportive lemurs are curiously named - they are amongst the least active of primates and their metabolism is about 50 per cent lower than would be expected in an animal of their size. Rather, their naming is said to reflect the position they adopt in defence, which resembles that of a boxer! They are nocturnal, largely solitary animals, very seldom venturing onto the ground. Members of the same sex defend territory against each other, in part using loud vocalisations, mixed with threat displays and chasing.
Question 9
Sportive lemurs are leaf eaters. What anatomical and physiological features would you expect as adaptations to such a diet?
Answer
You might expect an elongated large intestine (which is indeed the case) and a distinctive bacterial population to help digest the cellulose contained in the leaves.
The current distribution of the seven known species of sportive lemurs is shown in Figure 6b. The fact that some have geographical ranges that are so limited, often corresponding with the pattern of distribution of primary vegetation (shown in Figure 6a), reflects the vulnerability of these animals. In favoured locations, they occur in high population densities but they are still extensively hunted by humans; little is understood about many aspects of their biology.
5.4 Indris and sifakas
Both indris and sifakas are unusual amongst lemurs in that they are active largely by day. Leaves are their primary food. They have a specialised form of locomotion, best described as 'vertical clinging and leaping' and the leaps they can take, using their powerful legs, can be up to 10 m. DA describes the bounding movement of sifakas when they are compelled to come to ground. (If you go back to the previous video sequence that shows this strikingly white species - the so-called silky sifaka - bounding along on the ground, between 04:23 and 04:50.)
Sifakas coexist in families [p. 239]. Such groups are very variable in composition, with between one and 13 members. Females normally stay within one group but males move between groups, though the precise degree of promiscuity amongst males referred to in LoM p. 239 is uncertain. Male mammals that mate with several partners often have features that maximise the chances of successfully impregnating any one female. For example, such species normally have relatively large testes. Producing a large amount of sperm reflects a high degree of sperm competition prevalent in that species. (Sperm 'compete' in the sense that only one will fertilise an egg of a particular female.) In fact, sifakas have relatively small testes, which may reflect the fact that some other competitive system operates between males. It may be that some males achieve success by producing chemical signals from their scent glands, which in some way, at present not understood, suppresses reproductive function in rivals.
5.5 Ring-tailed lemurs
LoM p. 239 describes the life and habits of the ring-tailed lemur, drawing attention to what are commonly called their 'stink-fights' - a further example of the importance of smell in lemur society. But here the habit is prevalent in a species that is active by day and can spend as much as 40 per cent of its waking time on the ground. In fact, these animals seem equally at home on the ground and in the trees. Over time, some populations in Madagascar have become more ground-based than others, notably near the 2520m summit of the island's second highest mountain, where there is only sparse vegetation. You'll know something of the unique social system of ring-tailed lemurs [p. 239]. They live in large and complex groups and have a repertoire of calls, facial expressions, gestures and scent marking that allows extensive communication. Unlike all other primate groups, male ring-tailed lemurs are submissive to the adult females. When rival groups of ring-taileds meet, the dominant females take the lead. Females may also kill infants of their female rivals, whereas infanticide in lions and other mammals is usually performed only by mature males. The evolutionary reasons behind this unique instance of role reversal are not known.
5.6 Aye-ayes
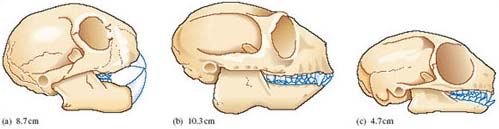
Perhaps with good reason, the aye-aye has been dubbed by some 'the strangest of all primates' and LoM provides some of the reasons [p. 243]. Little wonder that, as DA points out, the species was first classified as a rodent like them, it has powerful incisor teeth that grow continuously.
Question 10
Can you detect any other rodent-like feature of the aye-aye skull?
Answer
It has a diastema between the prominent incisors and the small premolars and molars.
The photograph in LoM p. 242 shows the spectacular elongated finger, which is the key to its ability to locate and catch the insect larvae contained within bark.
Question 11
With what tree-dwelling mammal, mentioned at the beginning of this course, does the aye-aye show a remarkable degree of convergence?
Answer
The striped possum (Section 1) which, as a marsupial, is taxonomically about as far removed from the primates as possible. This animal too has an elongated (middle) finger, and it too taps on bark. But it's uncertain if this habit aids location, as with the aye-aye, or whether the aim is more simply to disturb insects hidden underneath.
Recently, it's proved possible using captive aye-ayes to investigate what cues the animal uses to locate buried insect larvae, in an attempt to distinguish between the alternatives suggested in LoM p. 243.
Question 12
Aye-ayes have been found to excavate cavities that contain no prey. What does this suggest about what triggers excavation?
Answer
That any 'faint rustling' of the type that DA refers to is not the trigger that prompts the aye-aye (in DA's words) to turn 'to carpentry'.
Question 13
Perhaps then the alternative suggestion holds - that echoes from the tapping digit might reveal 'like a human doctor percussing a patient' where the wood is hollow. How might this suggestion be experimentally tested?
Answer
By filling the type of cavity inhabited by a larva, say with foam or gelatin, so that it is no longer hollow.
When this experiment was done, the aye-aye was no less eager to excavate; varying the density of the filling material (in ways likely to alter the reverberation of sound) made little difference to the aye-aye's behaviour. So, neither of the suggestions in LoM seems supported by these experiments. What seems more likely is that aye-ayes excavate when they detect by sound any interface or fragmentation in the underlying wood - what the researchers call a 'break in the integrity of the wood'. Maybe the fair proportion of false alarms this tactic is likely to produce is offset by the fact that many wood-boring larvae are large and nutritious, providing a substantial payoff when located. Furthermore, the number of unrewarding cracks beneath the surface of tropical wood is rather less than might be expected. In the dead wood that often represents the aye-aye's hunting ground, the high humidity of the rainforest means that cracking in wood is less frequent. But the bigger question of exactly to what extent the aye-aye uses the result of its tapping to construct a 'picture' of what lies beneath the surface, is unknown.
6 Reflection
Most of the activities that you have done so far are based on your understanding of single sections that you have just read. Activity 8 which follows, is different. It requires you to assemble and integrate information over the whole 'Life in the trees' topic and is likely to require some extra effort and thought. Integrating information from different sections of the course; is an important element in building your study skills. You're asked here to assemble evidence in support of a particular proposal and find other evidence that might cast some doubt on that same proposal. Critical evaluation of a statement, often from a number of perspectives, is an important writing skill in science and requires you to use facts to support your argument.
The final activity in this study period sets you a more demanding writing task that will require you to look back on this study period in its entirety, find some evidence and think of its significance.
Activity 8
A number of theories seek to explain how primates originated and what factors have shaped their early evolution. The oldest of them (the arboreal theory) states that primate characteristics evolved in the way they did as a consequence solely of selection pressure from living in trees.
(a) Write an account, of up to 200 words, that summarises the evidence in support of this proposal. You'll need to look at the list of primate characteristics in Section 3, identify those that are most closely linked with living in trees, gather together information about each from other parts of the text, and then structure your answer in a coherent way.
Answer
The hands and feet of primates are generally suited to the grasping that would be essential to living in trees. The thumb and/or big toe are angled outward (i.e. divergent) and in many species this digit is manoeuvrable, sometimes to the point of being opposable. The hands and feet, therefore, have a wide span and can grip, i.e. are prehensile. There are fleshy pads at the tips of the digits, and the possession of nails (rather than the claws likely to have been present in primate ancestors) may well be further adaptations that ensure effective movement through trees.
Effective vision in early tree-dwelling is likely to have been important. Primates have eyes that are forward-pointing and their fields of view overlap, providing stereoscopic vision, which is important in judging distances when moving through trees and for seizing prey (and perhaps fruit). The sense of smell is generally less important, though this sense can sometimes be significant, for example for nocturnal species and for species that inhabit dense vegetation. The characteristic structure of the muzzle of primates may reflect a downgrading of the sense of smell.
Activity 9
(b) The arboreal hypothesis is currently being re-evaluated as other facts that might have shaped early primate evolution are being considered. One approach has been to look at arboreal animals that are not primates and see if they have adaptations identical to or different from those of primates. Another has been to look for features of primates that are less readily explained as a consequence of living in trees - such as their omnivorous diet and the reduction in the sense of smell. Write about 200 words summarising evidence of this type that is useful for evaluating the arboreal hypothesis.
Answer
The tree-dwelling habit is well established in non-primates. The kinkajou is primate-like in some of its features, especially its monkey-like tail. Other features contrast with those of primates, notably claws. Indeed, in many species, claws can be very effective aids to climbing (e.g. sun-bear and tamandua). Many squirrels are also well adapted to tree-living and the lack of an opposable thumb and prehensile feet does not seem a strong evolutionary disadvantage, neither does the apparent lack of stereoscopic vision. A wide variety of adaptations are apparent in arboreal species, many of which are different from those evident in primates.
The leaf-, flower- and fruit-eating habits are widespread in primates, but the group as a whole is regarded as omnivorous. A number of primates have features of their digestion that are akin to true herbivores, e.g. bush-babies and some lemurs. But if access to leaves was such a strong feature in shaping primate evolution, it is perhaps surprising that a capacity to digest cellulose is not more widespread. It is less easy to see the diminished importance of smell in primates as linked with tree dwelling. In some tree-dwelling non-primates (e.g. the kinkajou), smell is significant; many primates (e.g. lemurs) seem to rely extensively on smell - of value in an environment where vision may be restricted.
If you found this activity difficult, don't be concerned - these are tricky arguments, even for professional biologists! My advice would be to read your answer and compare it with mine and think critically about the strengths and weaknesses of what you've written. Look back at the text of the course and make sure that you're able to find the relevant information, so that next time 'evidence gathering' of this type will be more manageable.
As the answer to part (b) of Activity 8 implies, evidence of this type is now prompting a reappraisal of the arboreal hypothesis. Maybe the development of primate-style vision and grasping hands and feet and the reduction of curved claws are adaptations linked with feeding on the fruits and insects on the low-growing shrubs, rather than high up in trees. The view at present is that while the tree-dwelling habit had an enormous influence on early primate evolution, other pressures and forces were at work in promoting the evolution of this group and shaping it into the diverse assemblage we see today.
Conclusion
This free course provided an introduction to studying Environment & Development. It took you through a series of exercises designed to develop your approach to study and learning at a distance, and helped to improve your confidence as an independent learner.
Acknowledgements
The content acknowledged below is Proprietary (see terms and conditions) and is used under licence.
Course image: Tjarko Busink in Flickr made available under Creative Commons Attribution-NonCommercial-ShareAlike 2.0 Licence.
Grateful acknowledgement is made to the following sources for permission to reproduce material in this course:
The content acknowledged below is Proprietary and is used under licence.
Figure 1 & 2 Macdonald, D. (ed.) (2001), The New Encyclopedia of Mammals, Oxford University Press;
Figure 3 & 4 Napier, J. R. (1970) A Handbook of Living Primates: Morphology, Ecology and Behaviour of Nonhuman Primates, Academic Press/Elsevier Science;
Figure 5 (a) Miyake, S. et al. (1991) Three-dimensional analysis of the peculiar arterial patterns of the extremities in Lorisidae: the rete mirabile, Primatology Today, Elsevier Science;
Figure 5 (b) Suckling, J. A., Suckling, E. E. and Walker, A. (1969) Suggested function of the vascular bundles in the limbs of Perodictus potto, Nature, vol. 221, January 25 1969. With permission of Nature (http://www.nature.com);
Figure 6 (a) Garbutt, N. (1999), Mammals of Madagascar, Pica Press;
Figure 7 Macdonald, D. (ed.) (2001), The New Encyclopedia of Mammals, Oxford University Press.
Don't miss out:
If reading this text has inspired you to learn more, you may be interested in joining the millions of people who discover our free learning resources and qualifications by visiting The Open University - www.open.edu/ openlearn/ free-courses