Studying mammals: The social climbers
Use 'Print preview' to check the number of pages and printer settings.
Print functionality varies between browsers.
Printable page generated Saturday, 20 April 2024, 3:31 AM
Studying mammals: The social climbers
Introduction
In this course we will focus on the Anthropoidea, a suborder of primates that includes monkeys, apes and humans. We will concentrate our attention here primarily on monkeys. Colour vision, a large brain and intelligence are of great importance in the lives of anthropoids, enabling them to eat foods inaccessible to many other animals and to exploit social situations. In this course, we will be looking at characteristics of primates that differ, or are enhanced, in anthropoids and discussing these attributes in relation to the evolution of the large anthropoid brain and the evolution of humans.
This is the ninth in a series of units about studying mammals. To get the most from these units, you will need access to a copy of The Life of Mammals (2002) by David Attenborough, BBC Books (ISBN 0563534230), and The Life of Mammals (2002) on DVD, which contains the associated series of ten BBC TV programmes. OpenLearn course S182_8 Studying mammals: life in the trees contains samples from the DVD set. You should begin each course by watching the relevant TV programme on the DVD and reading the corresponding chapter in The Life of Mammals. You will be asked to rewatch specific sequences from the programme as you work through the course.
This OpenLearn course provides a sample of level 1 study in Environment & Development
Learning outcomes
After studying this course, you should be able to:
identify the charactistics of primates and explain the main differences between the two suborders, prosimians and anthropoids
describe the detection of colour and estimation of distance in primates and explain the advantages of stereoscopic trichromatic vision
discuss the various types of communication seen in anthropoids and explain how playback experiments contribute to understanding vocal communication
compare and contrast adaptations in primates with adaptations in other mammals
compare and contrast how species reduce the risk of predation and avoid competition with other species.
1 The anthropoids
As you work through this course you will come across boxes, like this one, which give you advice about the study skills that you will be developing as you progress through the course. To avoid breaking up the flow of the text, they will usually appear at the start or end of the sections.
In this course you'll come across aspects of primate biology that are by their nature complex. You'll appreciate that primates are sophisticated mammals, and a good deal of detailed research work - a flavour of which is provided in what follows - has revealed just how subtle and intricate their biology is. Also, as with other areas of biology, our scientific understanding of primate biology is incomplete, so searching questions that may come to mind as you read often can't be easily answered - at least not without a good deal more study. You may need to read some parts of the text that follows several times, building up your understanding step-by-step. Referring back to earlier sections often helps, for example to become confident of the new terminology. Don't become over-concerned with every single detail of the information given - you should focus on the underlying ideas and concepts.
As well as the course text, you will be using The Life of Mammals book (LoM) and related The Life of Mammals DVDs, as described in the introduction to this course. Before you go any further, watch 'The Social Climbers' on the DVD and read LoM Chapter 9. Unless stated otherwise, all the page references you encounter in this course will be to LoM.
The social climbers of this course, the monkeys, are members of the order Primates. Some of the characteristics of primates are thumbs and big toes that are opposable to some degree, flattened fingernails instead of claws, sensitive finger pads, prehensile tails (but not in all species), dentition suited to an omnivorous diet, and stereoscopic vision.
The primates in S182_8 Studying mammals: life in the trees - the lorises, lemurs and bush-babies - are, using the convention in LoM, referred to as prosimians. In this course we will focus on the other suborder of primates, the Anthropoidea - monkeys, apes and humans. Some taxonomists favour the exclusion of tarsiers from the prosimian grouping, including them within the newly named haplorhine suborder, alongside monkeys, apes and humans. Following the approach of LoM, throughout this course I'll stick to the traditional distinction between prosimians and anthropoids. I'll confine my attention here primarily to monkeys; you can learn more about the anthropoid apes and about humans in S182_10 Studying mammals: food for thought.
Throughout the TV programme 'The Social Climbers', David Attenborough (DA) stresses the importance of colour vision and makes many references to the large brain and intelligence of monkeys, which enable them to eat foods inaccessible to many other animals and to exploit social situations. In this course, I will be looking at characteristics of primates that differ, or are enhanced, in anthropoids compared with prosimians, and discussing these attributes in relation to the evolution of the large anthropoid brain and the evolution of humans.
2 Who are the anthropoids?
As you know, the suborder Anthropoidea includes monkeys, apes and humans. You will be aware from reading LoM that taxonomists group monkeys according to the shape of their nose: Old World monkeys are catarrhines and New World monkeys are platyrrhines [p. 264]. In fact, apes and humans share many traits with Old World monkeys, so they too belong to the catarrhrines, whereas the New World monkeys are sufficiently distinct to be contained within a grouping of their own. DA also mentions other anatomical differences between catarrhines and platyrrhines, notably their dental formulae [p. 266].
Old World monkeys, such as baboons, mandrills, mangabeys, guenons, macaques, colobus and langurs, are found in Africa and Asia; they belong to a single family (Cercopithecidae). The marmosets, tamarins and capuchin-like monkeys of the New World are found in Central and South America; they comprise two related families. Apes (gibbons, orangutans, gorillas, chimpanzees and bonobos) and humans are the subject of course S182_10, so I will say little about them here. As DA points out in LoM p. 272, apart from humans, very few primates are found 'outside the tropics', i.e. the zone around the Equator between the Tropics of Cancer and Capricorn. The regions between the hot tropical zone and the cold polar regions have moderate temperatures and comprise the temperate zones.
Question 1
From your reading of LoM, identify monkey species that are found in the temperate zones and state whether they are Old or New World monkeys.
Answer
You may have cited Japanese macaques [p. 272], rhesus monkeys [p. 272], Barbary macaques [p. 274], and golden, Yunnan and Guizhou snub-nosed monkeys [pp. 266-267]. They are all species of Old World monkeys. (The Barbary macaques include the famous 'apes' of Gibraltar.)
Anthropoids are long-living animals that typically produce single offspring and have a lengthy juvenile period before they reach sexual maturity. Most anthropoid primates are omnivorous, but as they diversified to occupy different ecological niches, some species became more specialised. Some eat mainly fruit (the guenons and spider monkeys), or seeds (the sakis), or leaves (the colobus, langurs and howler monkeys), or even grasses (the geladas), with associated adaptations to their digestive systems and dentition. You will recall from the TV programme that bearded sakis can crack open seeds with their teeth, an ability that is described in more detail in LoM pp. 252-253. The degrees of specialisation, however, are minor compared with those found in other mammals, such as the panda, as all species are able to revert to other foods when their preferred food items are unavailable.
Question 2
From your reading of LoM, recall another plant item that is an important food for some monkeys and name the species that feed on it.
Answer
Exudates from trees that are referred to as gum. Gum is a sticky, rubbery substance, formed when sap oozing from a tree solidifies in air. Gum is a major food source for marmosets. It's also important in bush-babies.
Activity 1
Watch the TV programme from 10.48-14.58 and read LoM p. 258. Observe how marmosets use their claws to grip onto the tree-trunks and their teeth to gnaw the bark. Write a description, in about 200 words, of how marmosets use their claws and teeth to obtain food and explain why their methods of obtaining food differ from those of other monkeys.
Answer
Marmosets are tiny, enabling them to venture onto thin twigs that cannot support the weight of larger monkeys. Their light body enables them to walk along twigs without shaking and alerting their prey. A marmoset approaches its prey - an insect - stealthily, placing each foot slowly and carefully. Then it pounces, using its stereoscopic vision to judge the striking distance with great accuracy.
Monkeys use their hands and feet, and in some species their tails, to grasp branches as they climb in the trees. The hands and feet of marmosets are big enough only to grasp twigs, and rather than nails they have claws that dig into the bark of larger branches and tree trunks, giving a firm hold.
Although marmosets feed on insects, as do many other primates, their main diet is unusual. Marmosets feed on gum; their caecum is enlarged to aid digestion and they have chisel-like incisors that can gnaw holes in tree bark, causing sap to ooze out. The wounds heal, sealing in the sap, so every day marmosets need to gnaw open each wound. Constant opening and reopening of a wound forms a large ring of scar tissue around the wound, termed a sap pit. Generations of marmosets 'farm' the same tree, resulting in sap pits throughout the tree's height.
It is thought that marmosets adapted to eating gum because it is an important source of calcium. Calcium is required for growth and maintenance of bones and teeth. (Larger mammalian species may also eat eggshells and bones of small prey, which are even richer in calcium.)
Question 3
Think of an experiment to find out if it is likely that marmosets eat gum to obtain calcium.
Answer
You would need to find out if marmosets require more calcium than they obtain from foods other than gum. One method would be to offer marmosets two containers of drinking water, one containing plain water and the other containing calcium-enriched water, i.e. water that has additional calcium, and to monitor which one is drunk most often during a set period of time. If the calcium- enriched water is preferred, then the hypothesis that marmosets require additional calcium is supported.
In fact, this experiment has already been carried out. Researchers found that not only do marmosets preferentially drink calcium-enriched water but also lactating females show the greatest preference, suggesting that they need additional calcium for feeding their young, as indeed do most other mammals.
This experiment demonstrates that marmosets require calcium in their diet, but does it confirm the hypothesis that marmosets eat gum because it is a source of calcium? The results provide evidence that marmosets seek out sources of calcium. But for the hypothesis to be fully supported, we need to demonstrate that other foods in their diet, such as insects, are poor sources of calcium and that gum is a good source of calcium, indicating that individuals eating gum are more likely to obtain sufficient calcium than individuals that don't eat gum. Researchers measured the calcium content of different items in the diet and their measurements of insects and gum were very much along these lines. Using all this information together, we can be reasonably confident that the hypothesis is true: marmosets obtain the calcium they need by eating gum.
3 How do anthropoids differ from prosimians?
3.1 Diurnal living
In three particular respects, anthropoids have evolved features that have given them a competitive edge over other animals, especially prosimian primates. They have a diurnal instead of a nocturnal pattern of activity, they form all year round male-female relationships and live in social groups (which evolved differently and independently in lemurs), and they communicate extensively through gestures and vocalisation.
A diurnal pattern of activity means that, like us, nearly all anthropoid primates are awake during the day and asleep at night. Prosimians, the great majority of which are nocturnal, have adaptations for nocturnal living, such as large eyes, sensitive night vision with a resultant loss of colour vision, and a well developed sense of smell. Many also have large ears that can move independently of each other. In contrast to prosimians, the diurnally living anthropoids have evolved superior stereoscopic and colour vision with an associated reduction in the structures and brain processes related to smell.
3.2 Colour vision
DA stresses that colour vision is very important in primates, not only because colour is used 'in sexual displays' such as advertising a female's receptiveness to mating [p. 275], but also to identify ripe fruit [p.247] and to select nutritious leaves [p. 255]. This section discusses these points in more detail and explains how the visual system in primates is able to detect colour.
White light is composed of light of different wavelengths, from 300-800 nanometres (nm); 1 nm is one millionth of a millimetre. To understand colour vision you don't need to know anything about the physical properties of light, other than each band of wavelengths produces light of a particular colour (see Figure 1).
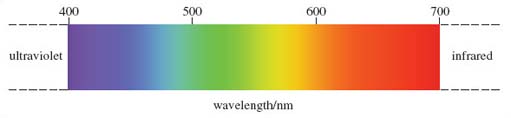
You may be familiar with the terms used to describe the shortest and the longest wavelengths, namely ultraviolet and infrared - over-exposure to ultraviolet light produces sunburn. Between these two extremes lie the wavelengths of the so-called visible spectrum, the light of various colours that humans can detect.
Light is detected by the retina at the back of the eye. In primates, as in the majority of mammals, light is detected by two types of so-called photoreceptor cell, namely rods and cones, which contain special light- sensitive compounds called photopigments. As stated by DA, cones are 'those elements in the retina that detect colour'.
So rods and cones are the receptor cells of the retina, and they are directly linked to nerve cells - more technically termed neurons. These neurons transmit information (in the form of tiny electrical signals) via the optic nerve to the visual cortex, the region of the brain that processes visual information. All neurons are interconnected; indeed, those with a specialised linking function are termed interneurons. They receive signals from one or more neurons and pass them to others, forming neural networks between the brain and the body. Each neuron has a long extension called an axon, which enables it to transmit a signal over a considerable distance before passing it to another neuron via a junction termed a synapse.
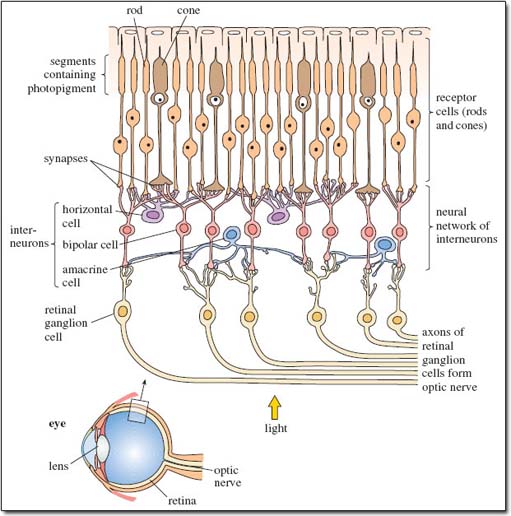
Figure 2 is a stylised diagram of the neural network of the retina showing the connections between the various cells. Spend a little time studying it. Locate the light-sensitive rods and cones (towards the top of the diagram) and the neural network of interneurons beneath them, which you'll appreciate is more complex in reality than shown here. Locate synapses, axons and various types of interneuron.
Light falling on the rods and cones produces signals that pass from receptor cells to interneurons, termed bipolar cells, and then to retinal ganglion cells whose axons form the optic nerve. There are two more types of interneuron in this network - locate them on Figure 2 - that form connections between different groups of rods and cones, such that signals from several receptor cells contribute to each signal entering the optic nerve. The communications that occur between these interneurons are too complex for discussion here, but this rich network contributes significantly to the sensitivity of primate vision. Rods are able to respond to low levels of light because a large number of adjacent rods (between 15 and 45) are linked to a single interneuron. Light only has to fall onto one of the rods for a signal to be transmitted to the brain. Not surprisingly, rods are the main or in some cases, as DA mentions on p. 257, the only component of the retina of nocturnal primates and are important for night vision in prosimians.
Cones need higher levels of light than rods to be activated and each cone is connected to between one and four neurons. In most species of New World monkeys, and indeed in all other non-primate mammals, cones contain two photopigments, one that responds to short-wavelength light and one that responds to medium-wavelength light; these photopigments may be found in the same cone or in separate cones. Electrical input from the activated cones is combined to produce colour images. This type of vision is termed dichromatic, in recognition of the two pigments.
Question 4
Dichromatic vision is not full colour vision, the type of vision that humans share with other anthropoid primates. Suggest why this is.
Answer
Because, unlike us, these animals cannot detect longer-wavelength light - they cannot distinguish red from other colours. (Refer back to Figure 1.)
Activity 2
You will recall from LoM and the TV programme that the only nocturnal anthropoids are owl monkeys. Watch the TV programme from 15.20-16.39 and reread LoM p. 257. Write a few sentences explaining why owl monkeys are thought to be descended from diurnal ancestors and how they compensate for poor vision.
Answer
Owl monkeys, like other nocturnal primates, have large eyes with retinas that contain many rods but no cones. Prosimians, however, also possess a tapetum lucidum, which reflects light towards the retina, greatly enhancing vision in low light. The lack of a tapetum in owl monkeys suggests that ancestral owl monkeys were diurnal, as are all other anthropoid species. In compensation for their poor vision, owl monkeys have an improved sense of smell compared with primates possessing colour vision. They mark trails through the branches, using a scent-impregnated tuft of hair that lies just beneath the tail.
How have you coped with the detailed anatomical diagram, Figure 2, showing the arrangement of cells in the retina at the back of the eye? Looking at the way different colours are used in the diagram should have helped you. You might find it useful to draw your own simplified version of Figure 2, perhaps as a series of boxes labelled 'rods and cones', 'interneurons', 'retinal ganglion cells' and 'optic nerve', linked by arrows to show the direction in which the nerve signals travel. Add some large arrows showing the direction of the light entering the eye - which, as the caption says, is not what you'd expect. Light reaching the rods and cones has had to travel through all the other cells. That gives you some idea of the transparency of cells, which are, in reality, packed very closely together, not arranged with lots of spaces between them as shown for clarity in this simplified figure.
3.3 Seeing red
Activity 3
Watch the TV programme from 20.37-24.06 and reread LoM pp. 247-248 and 255. Write a paragraph of about 120 words explaining why the inability to detect the colour red would disadvantage anthropoid primates. Try also to include any relevant information that you may have gained from studying other units in this series.
Answer
Although primates are omnivorous, leaves and fruit are important foods for many species. But many plants have evolved defence mechanisms to avoid predation. Most fruit is unpalatable and indigestible until the seeds inside are ready for dispersal. Similarly, newly emerging leaves may have high levels of poisons that can make primates unwell. As such leaves mature, the levels of poisons decrease but their relative toughness increases, making old leaves indigestible. Primates, therefore, need to distinguish ripe from unripe fruit and young leaves from old leaves. Recently emerged leaves on some tropical vegetation are red, becoming greener as they mature, and many ripe fruits are red or yellow, whereas unripe fruit are usually green. Primates that are unable to detect red would not be able to use these colour differences to select palatable foods.
Consequently, the gene mutation of an early catarrhine ancestor that led to the production of a third photopigment, which responds to red light, has been retained in Old World monkeys, apes and humans, presumably because the red-sensitive pigment confers a significant advantage.
Question 5
What term do you think biologists use to describe the visual system in Old World monkeys, apes and humans?
Answer
Trichromatic, because it involves three light-detecting pigments.
Trichromatic vision evolved convergently in only two genera of New World monkeys: howler monkeys and spider monkeys. As discussed in LoM pp. 248 and 255, any ancestral monkeys that were able to detect ripe fruit and red nutritious leaves more accurately had a competitive edge over other individuals.
Question 6
If this theory is correct, why is it surprising that trichromatic vision is rare in New World monkeys?
Answer
Because they also need to be able to detect ripe fruit and young leaves.
Recent research has perhaps solved this riddle. Researchers followed four anthropoid species feeding in the wild in Uganda. After the animals had left a feeding tree, the researchers compared the colours of leaves and fruit that had been dropped or nibbled on, with untouched leaves and fruit on the tree. Using meters that enabled them to mimic primate dichromatic and trichromatic vision, they found that dichromatic vision is sufficient to distinguish ripe fruit from unripe fruit, but not young leaves from old leaves, suggesting that detection of red leaves rather than ripe fruit may have been the main driving force behind the evolution of trichromatic vision. The selective pressure to detect red leaves may have been less in the New World because only 18-36% of young leaves flush red in New World vegetation, compared with 50-62% in Old World vegetation.
In diurnal mammals, cones and rods are scattered throughout the retina, with the concentration of cones increasing towards the middle. At the point where most incoming light is focused, there is a cone-rich area, the area centralis, which has a pit at its centre, the fovea (or yellow spot), that contains virtually no rods. These structures improve the visual image, making the detection of both predators and food items easier.
Food identification and detection of predators are not the only way in which primates use colour.
Question 7
From your studies to date, identify any additional ways in which anthropoid primates use colour.
Answer
Recognition of other individuals of the same species and selecting mates. You may recall the montage in the TV programme (25.36-25.46) showing the variety of markings amongst the guenons, or the badges of marmosets and tamarins mentioned in LoM p. 260. DA discusses mate selection several times in Chapter 9, reflecting the importance of colour in the ability of a male to attract females. For example, female uakaris prefer the males that have the reddest faces [p. 248] and female mandrills prefer males showing the brightest colours [p. 280]. Recent research has shown that female vervets prefer males possessing the brightest blue scrotums. Female of many species, such as baboons, signal their receptivity to mate by developing red sexual swellings [p. 275].
3.4 Stereoscopic vision
DA explains that ancestral monkeys had to 'decide whether a branch was within reach of a stretch or a jump - or beyond either' [p.248]. Individuals that were able to judge distances between themselves and branches more accurately had a competitive edge over other individuals, which led to the development in primates of stereoscopic vision. Stereoscopic vision evolved convergently in carnivores that judge distances to capture fast-moving prey.
The field of view of a single eye is termed the monocular field, which in most species is almost semicircular, i.e. the areas coloured pink and blue in Figure 3a. Many mammals, such as lagomorphs and ungulates, have an eye on each side of their head, i.e. laterally pointing eyes. In such an arrangement, the monocular field of view of the left eye has very little overlap with the monocular field of the right eye (Figure 3a), which maximises the field of view around an animal's head, enabling it to see predators approaching from almost any direction. Note that rabbits have unusually large monocular fields, giving them a circular field of view.
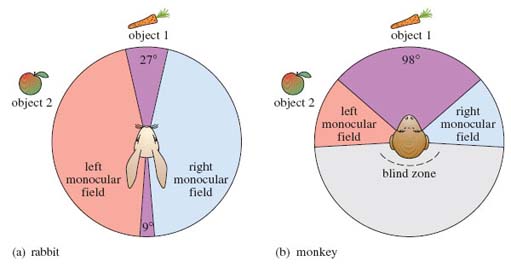
In contrast, primates have forward-pointing eyes, so the monocular field of the left eye overlaps considerably with that of the right eye (Figure 3b). This overlap reduces the total field of view so that all mammals with forward-pointing eyes have a blind zone - an area that their eyes cannot see; for example, you can't see behind you unless you move your head. But this region of overlapping fields is very important in primate vision. Objects within the area of overlap, such as object 1 in the figure, are viewed by both eyes, whereas objects outside this area are seen by either the left or the right eye but not both; for example, object 2 is seen only by the left eye. When the brain simultaneously receives information from both eyes about an object, complex neural processing occurs that builds up a 3-D image of the object. This is stereoscopic vision, which means literally solid vision, i.e. vision in depth, and hence enormously improves the ability to judge the distance between the viewer and the object. Try covering one eye and looking at a view or at a selection of objects on a table. The scene appears very 'flat'. Now look at the same scene with both eyes and note the illusion of depth.
The larger the field of stereoscopic vision, the greater the accuracy in judging distance. Primates, therefore, are able to judge distances with greater accuracy than herbivores. However, it is interesting to note that the large monocular fields in rabbits gives a small area of stereoscopic vision above and behind an individual's head, as well as in front, as you can see in Figure 3a.
All these developments in visual acuity increased the amount of processing that needed to be carried out by the brain and this is thought to have been the main driver of the large brain size that is found in living primates today (see Section 5).
As stereoscopic colour vision developed, so the sense of smell became less important. In the process, the nasal apparatus became less complex and most experts claim that this led to the shortening of the muzzle of anthropoids compared to other mammals. These changes to the muzzle allowed the muscles of the face to extend, giving mobility to the upper lip, enabling anthropoids today to make a wide variety of facial gestures. Facial and vocal communication are very important in anthropoid societies.
3.5 Communication
Compared with many other mammals, primates have a rich repertoire of communication skills, which I'll be looking at in more detail in this section.
Activity 4
Identify the instances of communication that you have observed in the TV programme or read about in LoM. Group these examples according to the type of communication involved, i.e. gestures, facial expression and vocal communication.
Answer
Gestures: You may recall vervet monkeys posturing to indicate alpha male status [p. 271] or baboons presenting hindquarters to indicate submission [p. 277-278].
Facial expressions: Many species use facial expressions, such as smacking the lips to show friendliness [p. 271] or raising the eyebrows or staring to indicate aggression (seen several times in the sequences of baboons and geladas). You may recall several instances of communication via facial expressions in the TV programme. A female tamarin uses her tongue to indicate that it is time for the males to carry the infants (17.18); a male and a female baboon flutter their eyelids and flash their teeth at each other to indicate they are both keen to sneak a mating whilst the alpha male is distracted (33.12); and a bachelor gelada gives an aggressive lip-curl towards a male that is established with a group of females, thereby signalling his readiness to fight for the females (43.42-46.33).
Vocal communication: You may recall vervet monkeys making alarm calls to indicate the detection of predators or strangers [p. 271], howler monkeys calling to defend their territory [p. 257], infant vervets calling for help [p. 271], or geladas involved in social chattering [p. 278].
3.5.1 Gestural communication
DA describes gestural communication as 'extremely eloquent' [p. 271] and such behaviour is well established in other mammals beside primates, for example cats and dogs. To attain this eloquence, gestural communication requires good visual acuity. In fact, gestures often rely on colour perception to enable individuals observing the gesture to respond accurately. For example, DA describes how De Brazza's guenons draw attention to their white-bearded dewlaps by shaking their heads when signalling aggression [p. 271], and dominant male vervet monkeys subdue subordinate males by flaunting their blue scrotums and red penises [p. 270]. The ability to use facial expressions has increased the complexity of social interactions between primates; therefore the brains of anthropoid primates also require a substantial amount of processing power to interpret gestures correctly.
3.5.2 Vocal communication
In order to be able to state that animals are communicating vocally with one another, scientists need to demonstrate that particular sounds made by one individual can be understood and acted upon by others.
Activity 5
You will be already aware that vervet monkeys make alarm calls in response to predators [p. 271]; as you saw in the TV programme, other species of monkey do this too. Watch the programme carefully from 26.02-30.30 and make comprehensive notes. You will also need these notes later. Write a few sentences to describe the monkeys' response to alarm calls.
Answer
The monkeys make specific calls in response to each predator. These calls are understood by other individuals, whose evoked behaviour reflects the nature of the threat. For example, on hearing the call reporting the sighting of an eagle, monkeys drop to the ground; on hearing the 'leopard' call, monkeys congregate near the 'caller', thus preventing the leopard from ambushing any isolated individual; on hearing a call indicating the presence of a snake, monkeys on the ground leap for the trees.
Question 8
Think of a way to test that the actions of the group are in response to a call and not to the actual presence of the predator. State the hypothesis that you will be testing.
Answer
One method is to use playback experiments. The alarm calls made in response to each type of predator (eagle, leopard and snake) are recorded. Each call is then played back in turn to the group, in the absence of any predators, using hidden speakers. The hypothesis being tested is: 'Each kind of alarm call alerts the group to the presence of a specific predator'.
Question 9
What do you predict would happen if this hypothesis was correct?
Answer
The group would respond to the recorded call as though an individual could actually see the predator.
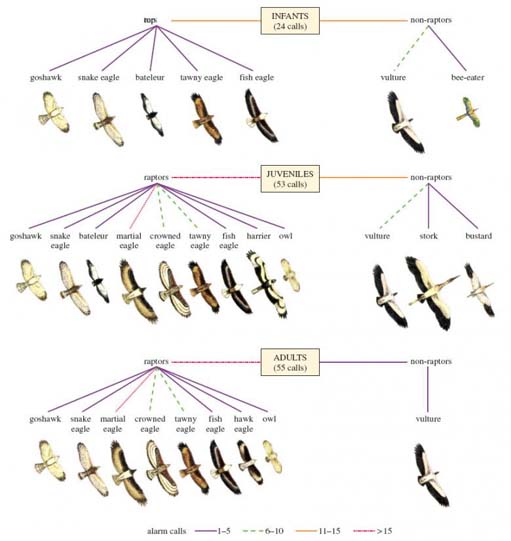
Exactly this response occurred when each type of call was played back to an experimental group of vervet monkeys, showing that vervet monkeys can make and interpret predator-specific calls. Infant vervet monkeys start making alarm calls when they are only a few days old. Of particular interest is how this behaviour develops over time. Figure 4 (below) offers a number of clues. It records the alarm calls made by infant, juvenile and adult vervets in response to various birds. Look at the data for infants at the top of the figure. Infant monkeys were observed calling in response to a bird on 24 separate occasions. The number of calls made in response to each of the species of bird shown were assigned to categories, shown by lines of different colours that are identified in the key at the bottom of the diagram. For example, the number of calls made by infants in response to a vulture is shown by a green line, indicating that between 6 and 10 calls were made.
Question 10
Study Figure 4 carefully. Write a few sentences to describe the development of alarm calling by vervets from infants to adults.
Answer
At first, infant vervets call in response to many species of bird, both raptors and non-raptors, but as they mature they become more and more proficient at recognising martial eagles (the main predator in that region). Note also that they make more calls in response to birds of similar size or similar wing pattern to martial eagles (e.g. crowned eagle) than to other birds.
How is this increase in proficiency brought about? Adult vervets can distinguish between calls made by juveniles and calls made by other adults. When juveniles call, the adults look around before reacting, presumably to check whether a predator has really been spotted; whereas adults react immediately to an adult call. Infants learn to make the same responses as their mother and they gradually learn to make calls only in response to a predator. The fact that infants can make recognisable calls only a few days after birth suggests that infant monkeys are born with an innate ability to communicate vocally, which is refined through learning. This type of innate ability may be one of the forerunners of our own language abilities. It is thought that human infants are born with an ability to separate human speech from other sounds, but the huge advances in human speech compared with monkey and ape vocalisations have involved many other factors, such as changes in the position of the larynx (voice-box) and developments in brain regions related to hearing and speech recognition.
Vervets also call in response to the presence of a rival group of monkeys. On sighting a rival group, individuals make 'wrr' calls to communicate to the rest of the group that rivals are approaching. As the rival group comes closer, the callers start making 'chutter' calls instead of 'wrr' calls. When members of the group hear 'chutter' calls, they move closer to the callers, ready to defend their territory. Playback experiments of the two types of call show that the group is responding to the specific call, not to the presence of the rival group alone. Playback experiments have also revealed some other insightful responses. When researchers play the 'chutter' call of a particular individual repeatedly in the absence of a rival group, the rest of the group eventually ignore it and carry on feeding or grooming, etc. Changing the recording to the 'chutter' call of a different individual causes the group to respond immediately again, but if it is changed to the 'wrr' call of the first individual, the group ignore that as well. Also, it was already known that vervet mothers respond rapidly to distress calls of their own infant, but playbacks of infant distress calls found that as a mother looked towards the sound of her infant's cries, the other females in the group responded to the sound by looking towards the mother.
Question 11
What does this behaviour tell us about the social abilities of vervets?
Answer
Vervets not only recognise the individuals in their group by voice alone but, remarkably, they can understand relationships between other individuals. Vervets are not unique in this respect, however, as many species of monkey show similar abilities.
4 Living in a society
4.1 Group size and collaboration
This section requires you to interpret the data presented in Figures 5 and 6. Figure 5 consists of two bar charts showing (a) the mean day range and (b) the weekly home range for a large and a small group of red colobus monkeys. They look rather similar, but check the labelling on the vertical axis. The day range is expressed in metres, while the weekly home range is given in hectares (ha). One hectare is an area of 10 000 square metres (m2). The text explains that the mean day range is a distance, whereas the weekly home range is an area. Remember always to check the units in which data are given. Notice too that these two diagrams show alternative ways of representing the same data; what is shown as an area (of feeding trees) on a map in Figure 6 is represented as the height of an individual bar on the bar chart in Figure 5b.
Primates can be found in variable-sized groups, from pairs up to groups of several hundred. Some of the driving forces behind group-living in other species of mammals are finding and defending food resources, and avoiding predation. For example, a pack of grey wolves can bring down a bison that may weigh nearly one tonne (1000 kg), prey that could not be captured by a lone wolf [p. 131]. For individuals that are prey for carnivores such as wolves, vigilance is vital. Herbivores such as zebra and impala herd together, so there is always at least one individual alert to danger and the chances of any one individual being singled out for attack are reduced.
Most researchers believe that overall size of a primate group is determined by food availability. Living in a group may enable members to defend their resources from other groups. However, as they forage together there may be less food available per individual compared to the amount that one individual could find by foraging alone. If food is readily available, the group can be large. For example, geladas feeding on savannah grasses live in very large communities of up to 800 individuals, but if a group has to travel to find food, it is likely that a large group has to travel further than a small group of the same species to satisfy their food requirements. This question was investigated in two groups of leaf-eating, red colobus monkeys living in the wild in Uganda. Researchers carried out what are called focal searches, concentrating on one individual in the group for a set time and then moving to another individual and then another, monitoring each for the same length of time. This procedure allows the movement of the whole group to be determined throughout the day. The researchers calculated the following measures:
the mean day range - the distance travelled by each group on average each day over a one-week period;
the weekly home range - the total area explored by each group in a week, for each of six consecutive weeks;
the total home range explored by each group during the six-week period.
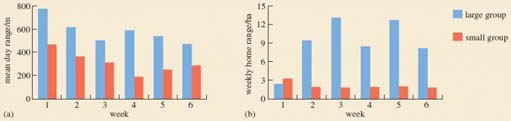
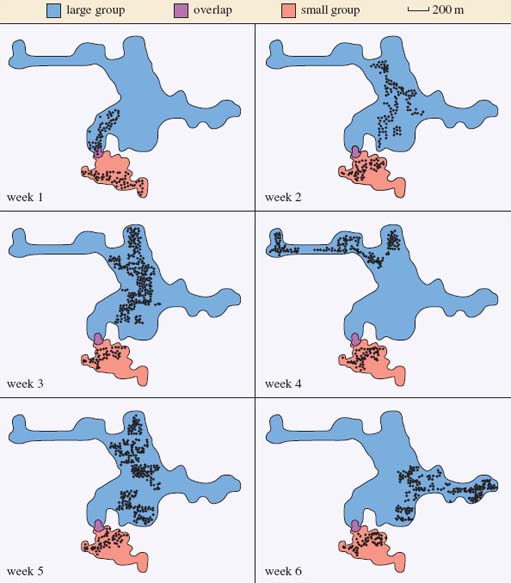
Activity 6
Look carefully at the data shown in Figures 5 and 6 above, including the information in the keys and in the captions. Note that the area of weekly home range is shown graphically in Figure 5b and as an area of feeding trees (dots) on the map of each total home range in Figure 6.
(a) What does Figure 5 tell you about the distance travelled by the two groups?
(b) You might well be bemused by results showing that leaf-eating monkeys are travelling up to 800 m a day. You will have already read that leaves are so nutritionally poor that other leaf eaters, howler monkeys, have little energy and tend not to travel far [p. 255]. Write four or five sentences suggesting why this statement may be true for howler monkeys but not true for red colobus monkeys [p. 266]. (You may wish to refer to LoM Chapter 4.)
Answer
(a) The mean day ranges and the weekly home ranges of the large group (in blue) are much larger than for the small group (in red), showing that the large group travelled considerably further than the small group. (Indeed, the small group never travelled as far in one day as the shortest distance travelled by the large group in a comparable time.)
(b) Both howler and red colobus monkeys have evolved digestive systems capable of supporting a mainly leaf-eating diet, but the adaptations differ between the two species. Howlers, like elephants and horses, are hindgut fermenters, possessing a caecum and an enlarged lower intestine that contains bacteria and protoctists capable of digesting cellulose, the main component of leaves. Red colobus, however, have cellulose-digesting microbes, similar to those found in howlers, in one or more compartments of a specialised stomach, an arrangement comparable to that of pseudo-ruminants. Although there are exceptions, the digestion of cellulose in the more anterior parts of the gut (as in the red colobus) is generally a very efficient process - more so than the comparable process in hindgut fermenters such as howlers. Such a difference may help explain why red colobus are able to expend more energy on travel.
It is worth noting that red colobus are still low energy users compared with other species of monkey; for example, fruit-eating spider monkeys travel much further than 1 km per day and may have a total home range of over 300 ha.
Question 12
Question: What can you infer about the weekly and total home ranges for both groups from Figures 5 and 6?
Answer
Each group forages within its own home range or territory although there is a small area in which both groups forage. The territory of the large group is much larger, in fact seven times larger, than the territory for the small group, which is surprising, given that the large group contained only twice as many individuals. The small group tended to forage in the same part of their territory from week to week, whereas the large group tended to move to a different area each week.
Question 13
Question: The data clearly demonstrate that the large group travels further per day across a larger area than the small group, but do they show that these differences are related to food availability?
Answer
The fact that the large group forages in a different part of its total range each week suggests that the food resources in one part of the forest are becoming depleted and the group has to move into another area to exploit new food resources. To confirm this suggestion, researchers need to investigate the food resources. If food availability is the same for each group, you would expect the large group to travel twice as far as the small group to gain the same amount of food per individual.
Question 14
Question: Suggest some factors that could be measured to compare the food resources available to the two groups.
Answer
You might suggest comparing the total number of feeding trees or the different species of feeding trees in the weekly ranges for each group. Or you might suggest determining the nutritious quality of the leaves (one estimate is the ratio of protein to cellulose, as protein is more nutritious than cellulose).
The other factor determining group size may be the need to avoid predators. In other mammals, such as savannah-living herbivores, as groups become larger, so vigilance can be shared between more individuals, reducing the risk of predation. Primate groups also tend to be larger in areas with high predation than in areas with lower predation, but what happens if the habitat cannot support large groups of particular species even though the risk of predation is high?
Activity 7
(a) From the TV programme, suggest how some monkey species reduce the risk of predation.
(b) Why might you find such behaviour surprising?
(c) Referring to your notes from Activity 5, write a few sentences to explain how monkeys in these associations reduce competition and improve cooperation.
Answer
(a) By forming mixed-species associations, in which different species, such as emperor and saddleback tamarins or sooty mangabeys, guenons, Dianas and colobus monkeys travel and forage together. Vigilance is increased because there are more individuals present than if the species were travelling and foraging separately. (See the sequences from 18.20-20.38 and 26.03-30.35.)
(b) You might suggest that this behaviour is surprising because normally you would expect different species of primate to compete for resources, or because such cooperation requires all species in the association to be able to respond to each other's alarm calls.
(c) Competition for food is usually avoided because each species occupies different levels within the canopy. In the Tai Forest, sooty mangabeys are on the ground, Diana monkeys are at the top of the canopy and several other species occupy the middle layers. Individuals in these associations are able to recognise and respond to alarm calls generated by any of the other species as well as their own.
In fact, these associations have given some fascinating insights into vocal communication in monkeys. As explained in LoM, 'the cat sat on the mat' is not the same as 'the mat sat on the cat' [p. 271], but until now, there has been no evidence of monkeys in the wild comprehending such syntactic rules.
Recent research, however, suggests that Diana monkeys foraging with Campbell's monkeys may be able to recognise the order of a series of calls. It was found that:
When Dianas hear a leopard or an eagle alarm call from a Campbell's, they give the corresponding alarm call of their own.
Male Campbell's monkeys also make a third type of call, a low-pitched boom.
If a Campbell's perceives a lesser threat, such as a falling tree, he utters two boom calls followed by a leopard call - a series of calls referred to as a boom-introduced alarm call.
When Dianas hear a boom-introduced alarm call they do not give leopard calls of their own.
Similarly, when booms were added experimentally to the eagle calls of a Campbell's and played back to Dianas, the Dianas made no eagle calls.
These responses suggest that boom calls alter the Diana monkeys' interpretation of subsequent Campbell's alarm calls, changing them from predator-specific calls to calls indicating a lesser threat. This conclusion is further supported by the observation that Dianas do respond to a played-back Diana call that is preceded by the boom calls of a Campbell's. This evidence is the first to suggest that the cognitive ability to generate and comprehend syntactic rules, albeit a very simple rule, evolved long before the emergence of human language.
Although these two factors (food availability and the avoidance of predators) influence the upper limit on group size in a particular environment, it is the rich diversity of internal relationships within anthropoid groups that is fascinating and has led to many studies of primates.
4.2 Social dynamics
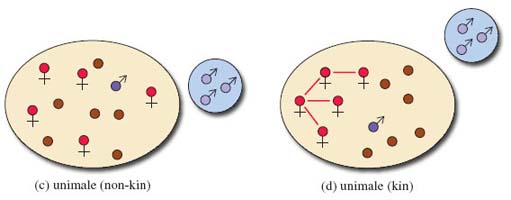
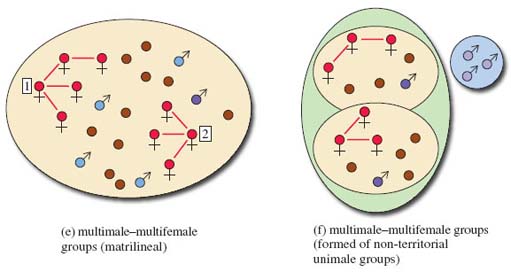
Yet another type of diagram is given in this section. Take a preliminary look at Figure 7, which is a way of illustrating the relationships between individual primates and groups of primates. Some specialist terminology is introduced and symbols and colours are used extensively. You will need to work slowly and carefully through the diagram to make sense of it all. Start by using the key to familiarise yourself with the symbols for a male (circle with an upward diagonal arrow) and for a female (circle on top of a cross). Younger primates of either sex are represented by just a circle. The coloured infill of the male symbol tells you something about the animal's status within the group. Then look at the make-up of each type of society and at the territory occupied. Look first at the simpler groupings shown in the diagram, (a), (b) and (c), to get used to the way territories are represented. You may find it helpful to make a few notes about each society in your own words, as you are referred to it from the text. For example, the first, labelled (a), consists of one adult male, one adult female and their young. Note that these diagrams do not include every individual present in a group but are schematic.
You will be aware from reading LoM that the majority of anthropoid primates, unlike most other group-living mammals, live in very complex societies that typically include adult males and females, subadults (adults that are not sexually active), juveniles and infants. The structure of the group differs between species. The major types of group composition are listed below.
one female and one male (monogamy, Figure 7a);
one female and two or more males (polygamy or more correctly polyandry, meaning more than one male, Figure 7b);
one male and several females (called unimale polygyny, or more often simply unimale), either non-kin (Figure 7c) or kin (Figure 7d);
several males and several females (multimale-multifemale); Figure 7e shows such a group, where the females are related (termed matrilineal).
There are more complex societies (Figure 7f and Figure 7g) that I'll describe later.

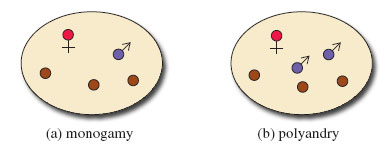
Activity 8
(a) Watch the TV programme from 31.00-48.19 and make notes on what you observe. (You'll be using these notes again in Activity 11.) The sequence illustrates the different social groupings of macaques, baboons and geladas. From your notes and your reading of LoM, identify which of these species form the following types of social groups: monogamous, polyandrous, unimale, multimale-multifemale, referring to Figure 7. (For this activity, ignore the issue of whether the females of a group are related, which I'll discuss later.)
(b) From the description in LoM p. 282, use the conventions in Figure 7 to draw a diagram representing a group of mandrills. Check your diagram with the answer below.
You might like to draw up a table at this point, showing the types of social grouping evident in the anthropoids described so far.
Answer
(a)
Marmosets form both monogamous and polyandrous groups [p. 264], as do tamarins, although only polyandrous groups are mentioned in the programme at (17.14).
Geladas [p. 278] and mandrills [p. 282] form unimale polygynous groups. In my terminology, this is what I refer to simply as unimale groups. (Don't be confused by the fact that DA calls these gelada and mandrill groups 'polygamous' [p. 282]; my preference is to avoid this less precise term - see the definitions above.) Uakari [p. 248], baboons and macaques form multimale-multifemale groups.
As you will recall from the programme, geladas form very large troops but these troops consist of smaller unimale groups that have banded together to avoid predation (Figure 7f).
(b) The mandrill society described in LoM consists of one male and up to 20 females with a few 'junior' males that have less developed colourings. This is represented below (Figure 8) as a multimale-multifemale (non-kin) group. You will see that it is similar to the multimale-multifemale (matrilineal) group shown in Figure 7e but the females are neither kin nor a matriline. (I have represented the junior males on my diagram using the symbol for subordinate males.)
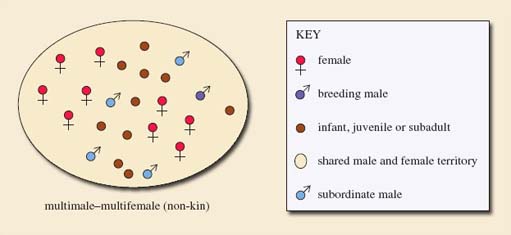
It is also important to note that in some species the social structure of the group depends also on the number of animals that can be supported by that particular habitat; a poor habitat may support a polyandrous or unimale group, whereas a richer habitat may support a multimale-multifemale group.
Most primate groups are usually formed around related (kin) or non-related (non-kin) groups of females that remain in the area of their birth. One or more males join a group to gain access to the females, and subadult males leave at maturity to find females of their own. In some species, such as red colobus, the groups form around male kin and the females disperse at maturity.
In primates, groups of males and females generally form year-long associations because males are needed to help the females raise their offspring, either through paternal care, as seen in marmosets and tamarins, or through defending the territory from other groups.
As I've mentioned, the simplest society consists of a monogamous pair and their offspring: the current infant and up to three subadults from previous litters (Figure 7a). But monogamy is rare among anthropoid primates, found only in marmosets, tamarins, siti monkeys and gibbons. As you will recall from LoM p. 264 and the TV programme, marmoset and tamarin females usually give birth to twins and the female needs both the male and the subadults to carry the infants so that she can devote some time to foraging. In marmosets and tamarins, these small family groups may include a second male (polyandry, Figure 7b) and when two mature males are present, the female mates with both.
Activity 9
Watch the TV sequence on tamarins (16.41-18.18) and reread LoM p. 264. In a sentence, write down why females are thought to mate with both males in polyandrous groups.
Answer
So that each male 'thinks' he has fathered the offspring and willingly stays to help rear them.
Just as with humans, paternity can be determined by DNA fingerprinting. This technique reveals that each male has sometimes fathered one of the twins; you may notice that LoM makes a contrary claim [p. 264], but I'm describing more recent research here. Without the assistance of both males and the older juveniles, the newborn twins are likely to die; thus there is survival advantage in this system of communal care.
Activity 10
You will recall from LoM that young female marmosets remain sexually inactive until they leave the group; the presence of their mother suppresses production of reproductive hormones so the females don't come into oestrus. (This is the stage when females are receptive and can conceive.) There is another species of mammal, the marmot, a squirrel-like rodent, in which young females remain with their mother despite being fully adult. Read Section 4 on the concept of fitness in S182_3 Studying mammals: chisellers and write a brief account, of about 150 words, to compare and contrast the situations in marmosets and in marmots. Include in your answer any benefits to the mother and the young females that arise from this behaviour.
Answer
Young female marmosets remain with their mother to carry younger siblings, allowing the mother to devote her energy to foraging. The presence of their mother suppresses hormone production, preventing young females from becoming sexually active. Young female marmosets benefit from this behaviour through the acquisition of good parenting skills, which may enhance their lifetime reproductive success by increasing the viability of their future offspring.
In contrast, young female marmots remain with their mother to provide warmth for younger siblings during hibernation. Young female marmots are sexually active but their mother subjects them to stressful behaviour that prevents pregnancies and causes abortions. The strategies of both mothers achieve the desired result, preventing additional young being born that might jeopardise the lives of current youngsters. In both species, young females enhance their own inclusive fitness by increasing the viability of their siblings, and mothers enhance their own lifetime reproductive success by increasing the likelihood that their offspring (subadult and infant) will survive to reproduce themselves.
In general, primate infants are particularly vulnerable to attack by their own species because they are born singly or as twins and are fed milk for a substantial period. Lactating females are unable to conceive, so when a male successfully ousts a resident male from a unimale group, he generally commits infanticide to bring the females back into reproductive condition. This behaviour is widespread among primates, possibly because a male may only have residency, and therefore the chance to reproduce, for a limited period, typically several years of a long life, although chacma baboons sustain residency on average for only seven months! Infanticide is also seen in multimale-multifemale groups when new males join. Although new males are not readily accepted (recall the difficulties faced by a new male baboon trying to join a baboon troop in the TV programme from 40.26-41.20), take-overs occur sufficiently often for females to have evolved counterstrategies that prevent infanticide. One strategy is paternity confusion; females mate with different males, so none 'knows' if he is the father and all the males help defend infants against new males. As mentioned in LoM [p. 275], female baboons signal their receptivity to mate by developing brightly coloured genitals; but in fact they can produce this coloration in response to a new male before they are actually ready to conceive, which protects their infants from infanticide.
4.3 Hierarchies within groups
Within multimale-multifemale groups of the type portrayed in Figure 7f, complex social relationships exist. These animals are 'forced together' to defend resources and to avoid predation, but competition for food and mates is substantially greater in large groups. This internal competition has led to males or females or both sexes forming kin or non-kin dominance hierarchies. In some groups, all the females are related (Figure 7d) and in this instance individuals inherit their rank from the mother - a so-called matrilineal dominance hierarchy. Some large groups consist of several matrilines (Figure 7e) and in these cases, all the females descended from one mother (i.e. of the same matriline) are dominant over another matriline, which in turn are dominant over a third, and so on.
As DA states in the TV programme, it pays to be 'high-born'. Females in the highest-ranking matriline will obtain the best food and most of the matings. You will recall that a female toque macaque in the programme was even able to remove food from the mouth of an older and bigger female because she outranked her. In years with low food availability, only the alpha male and alpha female (the most dominant male and female in the group) may produce any offspring. Individuals in male dominance hierarchies gain rank by fighting, and the alpha male is the strongest and fiercest male. Eventually, the alpha male is beaten by a younger male.
Question 15
Question: From reading LoM and watching the TV programme, describe how rank is maintained and how disputes between individuals are settled.
Answer
Generally a subordinate animal gives way to a higher-ranking individual. Subordinates forage at the least favoured positions and move position if a more dominant individual approaches, as is seen in vervets [p. 272]. They submissively present their hindquarters, or their chest in the case of gelada baboons [pp. 277-279], and the more dominant individual is appeased. Disputes do occur and they can escalate into a fight that may lead to injuries, but interestingly, once the dispute is settled, the individuals groom each other extensively. This reconciliatory behaviour is thought to help maintain the cohesion of the group.
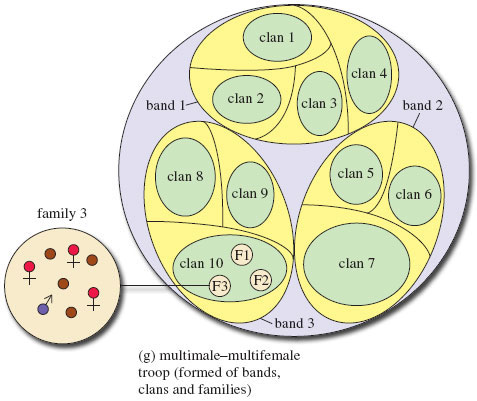
Some societies are very complex; for example, hamadryas baboon troops herd together at night, but during the day, as they forage within their home range, the troop separates into bands, clans and even families, depending on food availability (Figure 7g). In order to succeed within multimale-multifemale groups, such as those found in baboons and macaques, individuals form coalitions (or alliances) to help each other. Alliances are often kin-bonded but they also form between non-kin. Such alliances cut across the hierarchies and enhance an individual's survival and reproductive success. Both males and females may defend food resources and females may join together to prevent infanticide. A male may form a lasting friendship with a female, playing with, feeding and protecting her infant in order to gain matings next time she comes into oestrus. Males may form temporary coalitions so that the alpha male is distracted while one male sneaks a mating. This strategy can be successful; in one population of langurs the alpha male was found to have sired only 57% of the offspring, the remainder being fathered by his subordinates. Alliances occasionally help individuals move up the ranking. For instance, if a subordinate male aids the alpha male, the alpha male may reward him by increasing his rank. You will recall from the TV programme (34.00-34.34) alpha male toque macaques forming alliances with other males who guard the rest of the females while the alpha male mates. These non-sexual relationships are maintained through grooming. Within the group are several grooming cliques. Kin are more willing to groom each other than non-kin, and subordinates spend more time grooming higher-ranking individuals than vice versa. In times of crisis, grooming partners come to each other's aid. Grooming releases a 'pleasure hormone', beta-endorphin, that may help to reduce stress and tension within the group.
Question 16
Question: Why do you think kin are more willing to help each other than non-kin?
Answer
Because kin share genes with each other. For example, each individual has two parents, giving rise to the statement that an individual inherits half its genes from its mother and half from its father. Continuing this line of reasoning, a parent and its infant share 50% of their genes, as do siblings, because they have the same parents. Each individual has four grandparents, so grandparents and grandchildren share 25% of their genes. When an individual helps a relative, it is helping to maintain some of its own genes in the population, thereby increasing its own inclusive fitness.
As noted in LoM, primates also show tactical deception [p. 271]. On that occasion, by making a snake call, an individual 'stole' a tuber from its owner, who had just painstakingly uncovered it, causing the tuber owner to leap for cover and abandon the tuber. Juveniles also 'steal' food by uttering distress calls close to an adult that has a food item. The calls attract kin to the caller's aid, who, assuming the adult was attacking the juvenile, attack the adult. The adult drops the food and runs away and the juvenile eats it. Subordinates may also keep quiet when they see a predator, presumably because this behaviour makes it more likely that a higher-ranking animal will be caught.
This course is too short to discuss all the nuances and subtleties of the social life of primates, but this description should give you an indication of the importance of social interactions and the necessity for each individual to recognise relationships between not only themselves and another individual but between that individual and a third party.
Section 4 introduced a lot of terminology that was probably new to you. You'll appreciate that terms of this sort have to be devised by scientists to describe accurately the very different social groupings evident in primates, and it's important that you're aware of how this is done. I don't expect you to remember all these terms, but you do need to be able to apply them and you should appreciate the type of variation that exists in primate societies and the logic behind the use of such terms; so that when you are given information about primate social groupings, you can devise diagrams of this type using appropriate symbols.
5 The primate brain
5.1 The neocortex
Throughout LoM Chapter 9 and the TV programme, DA has referred to the large brain of monkeys and to their intelligence. Humans are the most intelligent of the anthropoid species, but evidence for non-human primate intelligence is growing.
Activity 11
Watch the TV programme from 01.26-09.14 and look back at the notes you made for Activity 8. Take this opportunity to look for examples of the issues raised in the previous section on primate societies. Summarise, in about 200 words, the points made concerning the evolution of the primate brain and the cited incidences of intelligence. You may also find it helpful to reread LoM pp. 248, 251 and 271.
Answer
The evolution of the anthropoid brain is thought to be related to the development of trichromatic vision, as the monkey brain is far larger than the prosimian brain. But other researchers believe that although vision was a contributing factor, it was the complexity of social living that was the main driving force behind the developing brain. Indeed, monkeys today show complex social behaviour, forming alliances to help maintain an alpha male's rank, forming friendships to gain matings or to be accepted into a group, and practising deceit to sneak matings when the alpha male is distracted. They maintain friendships through grooming, both actual and vocal, and infant care. Additional evidence demonstrating monkey intelligence comes from several sources. Rhesus macaques in Japan bathe in hot springs when the temperature falls. Capuchins rub themselves with piper leaves (which release chemicals that act as a natural insect repellent and antiseptic) when mosquitoes and skin infections are prevalent; capuchins have also developed a method for opening clams. Baboons can hunt flamingos. The striking thing about these behaviours is that they have been started by one individual and spread through imitation or direct teaching from one individual to another until the entire population is exhibiting that particular behaviour - so-called cultural transmission, which has its counterpart in many human interactions.
This final section will explore the evidence that scientists have obtained to support the claims that brain size in primates is related to colour vision and group size, and consider why such relationships help us to understand how the subsequent increases in brain size seen in apes and humans may have arisen.
Throughout the class Mammalia (and other vertebrates, for that matter) brain size increases with increasing body size; for instance, an elephant brain weighs approximately 8000 g, whereas a rat brain weighs less than 3 g (Figure 9a).

Question 17
Question: From your previous reading can you suggest some possible reasons for this relationship between brain size and body size?
Answer
The brain maintains all the functions of the body: growth, movement, repair, digestion, breathing, etc., the so-called somatic (relating to the body) processes - as well as learning and memory. As body size increases, animals require a larger brain to support and control these processes.
When scientists take body size into account, however, by calculating the ratio of brain size to body mass for each species (termed relative brain size) some species have larger brains than would be expected from their body sizes. But strikingly, primates have much larger brains for their body size than most other mammals (Figure 9b).
This finding intrigued researchers because brain tissue is known to be metabolically costly. Basal metabolic rate (BMR) is the rate of energy expenditure by an organism at rest at a non-stressful temperature. Scientists have also calculated the cerebral (brain) metabolic rate and shown that whereas other mammals use 2-6% of their BMR on brain maintenance, most primates use 9-14%. The most advanced primates, humans, use a staggering 20%. If the primate brain has evolved to be large in spite of these costs, the increases in brain tissue must confer important advantages.
Question 18
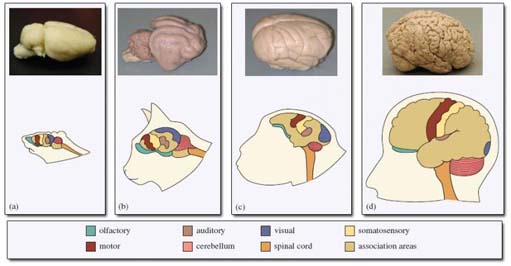
Question: What do you notice from the photographs in Figure 10 about the external appearance of the brain in primate species compared with other mammals?
Answer
The brain of primates is a more rounded shape compared with that of the other mammals. The outer layer of the brain is also more convoluted in primates.
This convoluted appearance is due to folds in a particular region of the brain, the neocortex, a thin layer of cells about 2 mm thick that covers the outer surface of the brain. (Neocortex is the term generally used for the cerebral cortex in primates.) When the ratio of the neocortex volume to the volume of the rest of the brain is compared across the primate order, it is clear that this ratio (termed relative neocortex size, or simply neocortex size) generally increases from the prosimians, through the monkeys to the great apes, and finally to humans. So why is the neocortex so important? You will already know from Section 3 that the evolution of colour vision led to an expansion in the visual cortex, the part of the brain that receives neuronal signals from the retina of each eye. The visual cortex is a region of the neocortex, so expansion in this area will contribute to increasing neocortex size. But colour vision is unlikely to be the only factor influencing the large brain size seen in primates; colour vision has evolved independently in fish, birds and amphibians, which do not have comparably large brains for their body sizes. Other regions of the neocortex receive input from other senses (Figure 10), such as an auditory (hearing) area, and a large part of the neocortex is involved in controlling movement of the body, i.e. motor control. The remaining parts of the neocortex are termed association areas, areas that are involved in higher thought processes, memory or creative thinking.
Question 19
Question: What is the most striking difference between the neocortices of the four species shown in Figure 10?
Answer
As the total size of the neocortex increases from rat to cat, through monkey to human, the association areas show a greater increase in size than the sensory and motor areas. This disproportionate change is interpreted as meaning that the major increase in primate brain size compared with other mammals is related to 'creative thinking'.
5.2 Social and ecological factors
Researchers think that the increase in creative thinking capacity was driven by group living because, in primates, increasing neocortex size is linked to increasing group size. We know that, even if there is not a strict hierarchy, monkeys are aware of the rank and kinship of each individual in their group, both in relation to themselves and in relation to each other. So as group size increases, each individual needs to keep track of a rapidly increasing number of relationships, which may be facilitated by the association areas of the neocortex. This relationship between group size and neocortex size is further supported because other aspects of social living are all related to neocortex size - factors such as the size of grooming cliques, the amount of tactical deception shown by individuals of the species, the amount of time juveniles spend playing and the length of the juvenile period, which could be regarded as the period for learning and socialisation.
But social factors are not the only aspects of primate life that are linked to neocortex size. Other researchers have shown that neocortex size is also linked to ecological factors such as the percentage of fruit in the diet and the size of the home range. As ripe fruiting trees are usually unevenly distributed throughout the home range, individuals need to move quickly from one food resource to another. Researchers suggest that individuals who were more able to hold a mind map of the locality in their head may have been more successful at finding food and this could be linked with an increase in neocortex size.
You have met several ways of presenting numerical data in this course. Figure 4 showed the number of alarm calls of vervet monkeys grouped into categories, 1-5 calls, 6-10 calls, etc., and then each category was represented by a different-coloured line. Bar charts were introduced in Figure 5, grouping data about the range of the red colobus monkeys. Figure 9a and b are also bar charts, with the length of each bar representing the absolute or the relative size of the brains of various mammals.
The graphs in Figure 11, used in Activity 12, are rather different. They are called scatter plots. In this example, the vertical axis (also called the y-axis) of each of them has a logarithmic scale. (You'll notice that Figure 11a has a logarithmic scale on the horizontal axis, i.e. the x-axis, too). In all three graphs, the x-axis represents the ratio of the neocortex volume to the volume of the rest of the brain. In general, scatter plots can be used to see if there is a relationship (also called a correlation) between the two variables plotted on the axes. Note that there are no units on the x-axis because ratios are relative measures.
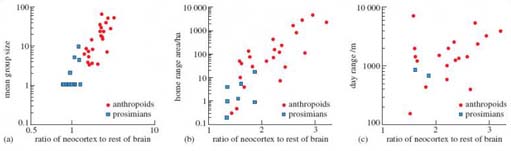
Activity 12
Study Figure 11 carefully, including the figure caption and the key provided. Can you decide at this point which of the three factors - mean group size, home range area and day range - is more likely to have an influence on the evolution of the neocortex? Write down your answer and then work through the explanation of Figure 11 that follows.
Figures 11a, b and c are scatter plots. Look at plot (a). Each symbol represents the mean value of group size plotted against the value of neocortex size for a particular anthropoid species. To see if a relationship between two variables exists, you need to look at the pattern of the scatter. Firstly you need to establish a 'line-of-best-fit' through the scatter of points so that the points are equally distributed on either side of the line.
Draw a line-of-best-fit through the points in Figures 11a, b and c. Now look at the way in which the points are distributed around each best-fit line. If the points lie close to the line, there is a stronger relationship between the two plotted variables than if the points lie further away from the line. When considering relationships shown on scatter plots, the steepness of the line also needs to be taken into account. A reasonably steep line (say, drawn at an angle of about 45 degrees), with minimal scatter, would mean that the values plotted on the vertical (y) axis change considerably for a small change in the values plotted on the horizontal (x) axis, showing a strong relationship. An almost horizontal line (nearly parallel to the x-axis) shows there is little change in y as values in x increase, meaning the relationship between the two variables is weak. Similarly, an almost vertical line (nearly parallel to the y-axis) shows that there is little change in x as y increases.
Question 20
Question: Look at Figure 11a, b, and c. Taking into consideration the scatter of points about your line-of-best-fit and the steepness of the line, can you order the plots by the strength of the relationships between the two variables plotted, starting with weakest?
Answer
You probably found it easy to put (c) into first place as the weakest relationship. Because the scatter is wide, it is difficult to decide where exactly to put the line through the points and the slope of the resulting line is quite shallow. You can fit a line more easily through (a) and (b), but deciding which relationship is the stronger is quite difficult.
To make comparisons between relationships easier, statisticians calculate a value called r (the symbol for a term called the correlation coefficient), which takes into account the position of the line, its slope and the spread of the points about the line and enables them to state whether the relationship is significant, i.e. 'Is it strong enough to be meaningful?'. The r values for (a), (b) and (c) are 0.87, 0.77 and 0.55; in the particular circumstances in which this test is being applied here, a value of 0.55 or above is taken as significant. Surprisingly the relationship between day range and neocortex size (c) is significant despite the large amount of scatter. (Strictly speaking, the relationship is between the logarithm of day range and neocortex size, but it's easier if we use the more convenient shorthand.) The relationship between group size and neocortex size (a) is slightly stronger than the relationship between total home range and neocortex size (b) although both relationships are significant. So the correct order from the weakest to the strongest is (c), (b), (a).
It's appropriate at this point to understand that this type of correlation can only measure the strength of a relationship between two variables. No correlation, no matter how strong, can prove that a change in one variable is causing the change in another. In this example, it's not possible to be certain that increasing social and ecological factors were the driving force behind increasing neocortex size. It's possible (though unlikely) that a third unidentified factor caused both variables to change hand-in-hand.
Figure 11a includes data for many primates, both solitary and group-living. When these data are looked at more closely, it's found that the neocortex size of solitary anthropoids is not greatly different from the neocortex size of prosimians that are solitary, whereas the neocortex size of group-living anthropoids is considerably larger than for solitary anthropoids and prosimians, thus providing further support for the theory.
But differences of opinion amongst researchers remain. As both group size and total home range size (and the other factors mentioned previously) correlate well with neocortex size, researchers are split; a few support the importance of ecological factors, but the majority believe that social factors are more important. Some very recent research, however, suggests that both sets of factors may have played an important role in the evolution of the primate brain. By collating from the literature all the examples of innovation, tool use and social learning in primates, researchers have shown that all three measures are correlated with neocortex size. Many of these examples of innovation, tool use and social learning are related to obtaining food, suggesting that both ecological and social factors have played a part in the evolution of the large primate brain that has led to the abilities of the most successful primate - humans.
Conclusion
DA opens the TV programme by stating that 'monkeys and apes have the richest social life of all mammals'. I have explored the importance of colour vision in the interactions between individuals and discussed how gestural and vocal communication add considerably to this richness. But ultimately, it is their ability to be innovative, to discover new ways of obtaining foods, to learn from one another (so-called cultural learning), to form friendships, alliances and coalitions between individuals, and to practice tactical deception that have fascinated and continue to fascinate primatologists as they strive to understand anthropoid societies and to seek out forerunners of the abilities and sociability of humans. If you get an opportunity to watch a group of monkeys, observe them closely and see if you can recognise some of the aspects of their social life that we have discussed in this course.
Acknowledgements
The content acknowledged below is Proprietary (see terms and conditions) and is used under licence. This content is made available under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 Licence
Course image: Harald in Flickr made available under Creative Commons Attribution-NonCommercial-ShareAlike 2.0 Licence.
Grateful acknowledgement is made to the following sources for permission to reproduce material in this course:
The content acknowledged below is Proprietary and is used under licence.
Figure 2 Frisby, J. P. (1980) Seeing, Illusion, Brain and Mind, Oxford University Press;
Figure 3 Bradbury, J. W. and Vehrencamp, S.L. (1998) Principles of Animal Communication, Sinauer Associates, Inc.;
Figure 4 & 10 (bottom) Gould, J. L. and Gould, C. G. (1999) The Animal Mind, Scientific American Inc.;
Figure 6 Gillespie, T. R. and Chapman, C. A. (2001) Determinants of group size in the red colobus monkey (Procolobus badus): an evaluation of the generality of the ecological-constraints model, Behavioural Ecology and Sociobiology, vol. 50, no. 4, p. 333, fig. 1A-F, copyright Springer-Verlag 2001;
Figure 9 Macdonald, D. (ed.) (2001), The New Encyclopedia of Mammals, Oxford University Press;
Figure 10 (top) Sourced from http://serendip.brynmawr.edu;
Figure 11 Dunbar, R. I. M. (1989) Neocortex size as a constraint on group size in primates, Journal of Human Evolution, vol. 20, pp. 478 and 484, copyright 1992, with permission from Elsevier.
Don't miss out:
If reading this text has inspired you to learn more, you may be interested in joining the millions of people who discover our free learning resources and qualifications by visiting The Open University - www.open.edu/ openlearn/ free-courses