Studying mammals: The insect hunters
Use 'Print preview' to check the number of pages and printer settings.
Print functionality varies between browsers.
Printable page generated Thursday, 30 April 2026, 8:22 AM
Studying mammals: The insect hunters
Introduction
Sixty-five million years ago, animal and plant life were very different from nowadays, but there were rat-sized placental mammals living successfully on the ground. They were insect eaters, i.e. insectivores, feeding on the vast numbers of insects and other invertebrates living in soil, leaf litter and low-lying vegetation. Insectivore means 'insect eater', and in this course we will explore the world of insect-eating mammals, classified together on the basis of a reasonably close evolutionary relationship.
This is the second in a series of units about studying mammals. To get the most from these units, you will need access to a copy of The Life of Mammals (2002) by David Attenborough, BBC Books (ISBN 0563534230), and The Life of Mammals (2002) on DVD, which contains the associated series of ten BBC TV programmes. OpenLearn course S182_8 Studying mammals: life in the trees contains samples from the DVD set. You should begin each course by watching the relevant TV programme on the DVD and reading the corresponding chapter in The Life of Mammals. You will be asked to rewatch specific sequences from the programme as you work through the course.
This OpenLearn course provides a sample of level 1 study in Environment & Development
Learning outcomes
After studying this course, you should be able to:
describe the lifestyle of a variety of insect eaters, from four orders
give examples of adaptations linked to feeding in insect eaters
explain the limited extent to which insectivores can be regarded as ‘primitive’
characterise typical adult mammalian dentition and understand dental formulae
recognise teleology and write down accounts of evolution that do not assume purpose or direction.
1 Meeting the insect eaters
As you work through this course you will come across boxes, like this one, which give you advice about the study skills that you will be developing as you progress through the course. To avoid breaking up the flow of the text, they will usually appear at the start or end of the sections.
As well as the course text, you will be using The Life of Mammals book (LoM) and related The Life of Mammals DVDs, as described in the introduction to this course. Before you go any further, watch 'The Insect Hunters' on the DVD and read LoM Chapter 2. Unless stated otherwise, all the page references you encounter in this course will be to LoM. When you have watched the TV programme and read LoM Chapter 2, you should have a good 'feel' for the diversity of insect eaters and the range of niches they occupy.
Chapter 2 of LoM begins by describing conditions within a super-continent in the Northern Hemisphere 65 million years ago, during the period of the geological past called the late Cretaceous. As author David Attenborough (DA) points out, both animal and plant life in the late Cretaceous were very different from nowadays. The dense forests were rich in ancient types of coniferous tree, and luxuriant tree ferns and cycads - the latter looking a bit like giant pineapples, with crowns of extensive fronds. Dinosaurs, including large predators, were diverse and abundant; nevertheless, rat-sized placental mammals were living successfully on the ground. They were insect eaters, i.e. insectivores, feeding on the vast numbers of insects and other invertebrates living in soil, leaf litter and low-lying vegetation. Available fossil evidence suggests that placental mammals had diverged from marsupials more than 90 million years ago, during the middle of the Cretaceous. You can think of this divergence as an early branch point in an evolutionary tree, where the two branches (placental and marsupial) sprang from a common ancestral type, each branch further subdividing over geological time into the wonderful array of types described in LoM.
In everyday language, insectivore means 'insect eater', but in LoM and in this course its meaning is much more precise. Insectivores are insect-eating mammals, classified together on the basis of a reasonably close evolutionary relationship. So, for our purposes, an insectivore is a member of the order Insectivora. But as DA points out (and the photograph on p. 39 very convincingly demonstrates), not all insectivores restrict themselves to a diet of insects; it is more accurate to think of insects as a significant proportion of their diet. Insectivores are defined on anatomical grounds, including their small size, their long and mobile snouts and their plantigrade habit, i.e. they walk on the soles of their feet with the heels of their hindlegs on the ground.
Insectivores are often said to be the most primitive of placental mammals, though you may know from S182_1 Studying mammals: a winning design (the first course in OpenLearn's Studying mammals series) that the whole notion of primitiveness is contentious. It is generally agreed that modern-day insectivores most closely resemble, in both their structure and their way of life, those ancestral mammals from which all modern mammals are thought to have evolved. But not all modern-day insectivores are primitive. As you'll see in this course, many insectivores have evolved specialised structures and behaviours that to different degrees disguise an underlying primitiveness.
SAQ 1
What group of living mammals does DA suggest resembles the earliest insectivorous mammals?
Answer
The earliest mammalian insectivores are described as small and shrew-like [p. 38].
Shrews occupy the same niche as the earliest insectivorous mammals - small and ground-dwelling, feeding on insects in leaf litter and undergrowth. The term 'niche' is used to describe an animal's role within its particular environment. It reflects various characteristics, such as habitat range, how the animal feeds, how it captures its food, its environmental requirements, the food it eats and what are its predators. A niche is, therefore, a description of lifestyle within a particular ecosystem - a term that means a community of living organisms, including different species of plants, animals and fungi, which interact with one another, and with their physical environment. You'll learn more about ecosystems if you work through the remaining units in this series.
A variety of orders of placental mammals exist. Taxonomists argue a great deal about the evolutionary origins of these different groups and how the taxonomic boundaries ought to be drawn, so the exact number of mammalian orders varies from 19 to 27, depending on whose scheme you follow. But LoM focuses largely on members of just four orders of largely insect-eating mammals: Insectivora, Chiroptera, Pholidota and Xenartha. It isn't appropriate here to describe the defining features of each of these four orders in detail, but we can gain an impression of the diversity of insect-eating mammals from a brief exploration of various niches that they occupy, building on the coverage in LoM.
The order Insectivora comprises a very diverse range of mammals, including shrews, desmans, moles, tenrecs and hedgehogs - which LoM describes in that sequence. (You have the option of rereading pp. 38-44 and pp. 51-54 to refresh your memory.) Other insectivores are the golden moles, which I'll mention later; these animals are wonderfully filmed in the TV programme as 'they swim through sand'. Another group of insect eaters are the elephant shrews. You might recall these animals from the TV programme looking like large versions of true shrews, with their 'path-tidying habit' captured on surveillance camera. Many taxonomists now argue that elephant shrews are sufficiently different from insectivores (and closer to rabbits and hares) to warrant being assigned a separate order, all to themselves.
The long-tailed shrew tenrec [p. 52] also resembles a large shrew, and occupies the same niche, foraging in undergrowth and leaf litter for insects and other invertebrates. As a group, tenrecs (and their close relatives the otter shrews) are very diverse in appearance - a good many are aquatic. The family as a whole displays features that, to some degree, resemble those of ancestral mammals. For example, their body temperature is low and varies throughout a 24-hour period. There is also a cloaca - the common opening for the digestive and urinogenital systems - that is also present in non-mammals such as lizards.
Hedgehogs eat earthworms, slugs, eggs and frogs, in addition to insects. They, of course, are considerably larger than shrews and cannot always hide in undergrowth when foraging, so their niche is not quite the same as that of shrews. DA refers to hedgehogs 'fossicking' around the garden [p. 53] - a delightful term but one with no precise biological meaning, other than the type of 'rummaging around' feeding behaviour evident under DA's gaze in the TV programme.
Other insectivores occupy niches characterised by preying on insects that live in water and soil.
SAQ 2
From LoM and the associated TV programme list insectivores that forage for insects (a) underwater, and (b) underground.
Answer
(a) Desmans, star-nosed moles and water shrews dive underwater to forage for insects. (b) Moles and golden moles occupy burrowing niches, hunting insects and other invertebrates, such as worms, below the ground.
Bats, grouped in the order Chiroptera, have highly distinctive faces, sharp teeth and furry bodies; their defining feature is possession of wings. In the TV programme DA illustrates the huge range of bat diversity, showing that most bats feed on insects; a few feed on fish; some tropical species feed on fruit, and a few feed on nectar, frogs or mammalian blood. Most insect-eating species are nocturnal, preying on night-flying insects - a very different niche from that occupied by shrews. LoM suggests that bats evolved from a shrew-like ancestor early in the evolutionary history of the Insectivora [p. 44], and highlights the oldest known bat fossil, dated at 50 million years old. You'll probably recall the spectacular film sequences of the New Zealand short-tailed bat in the TV programme (at 44.16), which demonstrate its surprising agility on the ground, helped by a unique ability to tuck its wings tightly against its body. These bats catch prey in the air (using echolocation as described on p. 46), but rely upon hearing and sensitivity to smell when hunting in the leaf litter, which they do in a remarkably 'shrew-like' way.
Armadillos and anteaters, grouped in the order Xenartha, and pangolins, order Pholidota, occupy the 'ant-eating' niche, in which animals break into nests of colonial insect species, and eat large numbers during one hunting foray. In the TV programme, DA demonstrates evidence that shows that pangolins and anteaters have persisted unchanged for many millions of years. Fossil anteaters, tamanduas and pangolins, dated at 50 million years old, were found in the Messel Shales in Germany, and resemble species living today in South America and Africa. The TV sequence of a fossil pangolin 'coming to life' on the museum bench as an African pangolin is memorable (see the programme at 27.22), with just a glimpse of the unique 'shuffling gait' you saw in living specimens.
You have just met a lot of detailed information about the classification of insect eaters. You might find it useful to summarise this material for yourself in a notebook, so that you can refer to it as you progress through this course. You could copy down the four orders and list the names of the species mentioned in each of them. You can add extra species to your lists as you meet them. Similarly, as you read on through Section 2, you might like to make another list of the different sorts of teeth that are found in mammals, with their distinctive features. Teeth will be mentioned in several of the later units so, again, this list may be useful for reference. Whenever you study a section, consider whether it might be useful to summarise any of the information in a list of this type.
2 How insect eaters obtain their food
2.1 Mammalian dentition
Insects are generally very small animals. Many kinds are hard work to collect and not very nutritious because a high proportion of their mass is a protective and indigestible outer layer, called cuticle. Insectivorous mammals need to eat large numbers of insects to fulfil their energy requirements.
Insect eaters have diverse ways of catching and dealing with their prey; teeth play a crucial role. Indeed, teeth are of such enormous significance to mammalian diets in general (and are so readily preserved by fossilisation) that they feature extensively in this series of units as a whole. A general introduction to teeth is useful at this point.
Mammals have several kinds of teeth that operate together as a set, of which there are only two - the 'milk' set that emerges just before weaning, and the permanent set that replaces it usually before sexual maturity. Because the young are fed on milk for some time after birth, the emergence of the teeth is delayed until the jaws have grown large enough to accommodate them. Consequently, teeth in the upper and lower jaws grow to be opposite each other. This matching, together with the tough hard materials and complex structure of each tooth, are crucial to their role as precision tools in feeding. In fact, the possession of differentiated and exactly opposing teeth is a defining character of mammals. The downside is that, although reptiles such as snakes and crocodiles can replace damaged or diseased teeth throughout life, mammals have a fixed number of teeth and, as many of us know to our cost, defective teeth are not replaced.
At the front of the jaw there are incisors, used for grasping and biting food. Canine teeth behind the incisors are pointed, and often sharp, but may be small and poorly developed in animals that are not flesh eaters and do not use their teeth for defence and in threat displays. Premolars and molars behind the canines have several roots and are usually broad and ridged. These teeth are the main grinding surface for chewing food, a uniquely mammalian ability that entails complex movements of the tongue, cheeks and jaws. Sheep, cattle, deer and their relatives chew by side-to-side movements of the jaw, while rats, squirrels, guinea-pigs, etc. grind their food by sliding the lower jaw forwards and backwards. Other mammals including ourselves use combinations of these movements. The numbers of each type of tooth in a mammal can be shown in shorthand by the dental formula, of which you see examples later. Such a formula describes the numbers of incisors (I), canines (C), premolars (P) and molars (M) on one side of the upper and lower jaws. For example, if a mammal were to have three premolars on one side of its upper jaw and two premolars on one side of its lower jaw, this would be abbreviated to P3/2. The milk set, which develops before the jaw is fully grown, includes premolars but not molars. The milk set may be partially or entirely eliminated in some kinds of mammals, and some, notably the great whales, are completely toothless.
A typical mammalian tooth is made up of three regions: the crown, the neck and the root (Figure 1). The visible part of the tooth that emerges from the gum is the crown; the neck of the tooth is surrounded by the gum. The tooth is anchored in the gum by the root, and a layer of bone-like tissue, called cement, fixes the root to the jaw bone. In most mammals, the permanent teeth eventually stop growing and their blood supply is reduced, at which point the teeth are described as 'rooted'.
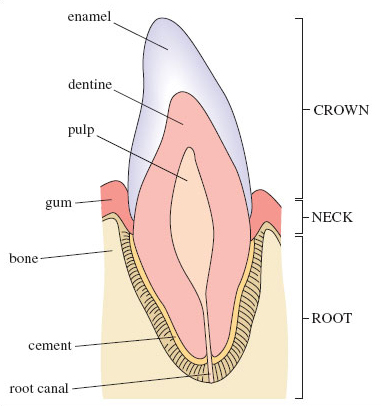
In vertical cross-section, the tooth can be seen to consist of three layers. The outermost layer of the crown is hard enamel, made of mainly calcium phosphate with a small proportion of calcium fluoride. The enamel resists damage from hard foods and protects the inner layers. Most of the tooth is made up of dentine, which is similar to bone in structure, made up of a fibrous framework, (rather like the mish-mash of coconut fibres in a doormat, but on a microscopic scale) in which are embedded crystals of calcium phosphate. The central pulp cavity contains blood vessels and nerves and becomes almost closed off at maturity.
Members of the Insectivora have numerous (up to 44) small teeth. Look at Figure 2a, which shows the skull and lower jaw of the hedgehog.
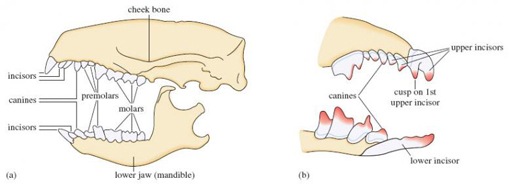
SAQ 3
How many incisors, canines, premolars and molars are there in the upper jaw of the hedgehog? Express these values using the shorthand given above.
Answer
There are three incisors (I3), one canine (C1), three premolars (P3) and three molars (M3).
Counting the corresponding numbers in the lower jaw of the hedgehog gives: I2, C1, P2, M3. Bear in mind that the dental formula gives, for each type of tooth, the ratio of the number in one side of the upper jaw to the number in one side of the lower jaw. The dental formula of the hedgehog is thus I3/2, C1/1, P3/2, M3/3, and the hedgehog has a total of 36 teeth.
SAQ 4
The pygmy shrew has three incisors, one canine, three premolars and three molars in one side of the upper jaw, and one incisor, one canine, one premolar and three molars in one side of the lower jaw. What is its dental formula and how many teeth does it have in total?
Answer
The dental formula of the pygmy shrew is I3/1, C1/1, P3/1, M3/3, giving 16 teeth on each side and so a total of 32 teeth. (Figure 2b shows just the pygmy shrew's teeth nearer the front of each jaw.)
2.2 Feeding techniques
In Activity 1, below, you are asked to make notes from a TV sequence and then select some of the information from your notes and combine it with some from LoM into a single short piece of writing. Check the wording carefully to find out exactly what you need to do. Use the LoM index to locate the information you need from there. You should aim to write a few clear sentences on the topic. When you have written your answer, compare it critically with the one given. In what ways does it differ? Have you included more facts, or missed some out? Have you used more words to say the same things? To write scientifically means to write very concisely and it may take you quite a lot more practice to write in this way.
Activity 1
Watch the TV programme on the DVD from 00.32-07.05 and make brief notes on the techniques used by pygmy shrews and water shrews for locating their food. From your notes and your reading of LoM, briefly describe how the pygmy shrew searches for insects. You will use your notes on water shrews in Activity 2.
Discussion
Pygmy shrews forage frantically for most of the time they are active, detecting ground-living insects by sound or scent. When searching for food, the pygmy shrew runs rapidly over the ground, digging vigorously through undergrowth and leaf litter with its front feet and probing with its snout. (I also noted that the small eyes of shrews provide poor vision, but their sense of smell is well developed and the whiskers are very sensitive to touch.)
The shrew's technique for searching for insects is very efficient. Researchers buried insects up to 12 cm deep in soil, grass and moss in an enclosure, and within 15 minutes a shrew had captured and eaten one earthworm, a cranefly (daddy-longlegs) larva, a large caterpillar and a slug. When a shrew finds an insect, it grabs it with the front incisors, which are long and pointed; the pair of first upper incisors have an extra cusp projecting at the base (Figure 2b). The shrew bites off the insect's head, holding the insect with the front paws, and eats it from the head down, biting off pieces with the incisors. As the food passes back towards the throat it is chewed by the molars, which have sharp surfaces. It has been estimated that a common shrew has to find and eat about 100 maggot-sized prey every 24 hours. This requirement is less of a problem than it sounds; there are up to 1000 insects per square metre on vegetation and in soil in scrub grassland.
Like shrews, hedgehogs spend most of their active time searching for food, but less frantically so. They forage at night and wander around walking quite slowly, sniffing and listening for prey. When a prey animal is detected by its scent, or by the noise of its movement in leaf litter, the hedgehog darts forwards or sideways to grab it. Hedgehogs, of course, grow to a much bigger size than shrews, and a large individual can be up 20 cm long.
I've already mentioned the hedgehog's varied tastes, a point echoed in LoM [p. 53]. Eggs and prey ranging from lizards to earthworms contain a wider range of nutrients than insects; for example, iron (needed for blood) and calcium (the major component of bones and teeth) are available from such prey. This is why DA links the larger body size of hedgehogs to the broadening of their diet [p. 54] - though at this point you're probably able to think of specialised insect eaters considerably bigger than a hedgehog (Section 3.4).
As you saw in Figure 2a, hedgehogs have 36 relatively small teeth. The incisors are pointed and project forwards, the canines are small and the molars have well developed W-shaped surfaces. Hedgehogs use their incisors to grab prey and may use the body and front paws to hold down larger prey. Large beetles are grasped by their side or hind-end by the incisors and crushed by the molars. When hedgehogs eat vertebrate carrion, they gnaw at the flesh rather than slashing it like true carnivores - their teeth are relatively small and not sharp. They take eggs from birds' nests on the ground, and also chicks, which they hold down with the front paws and eat alive, rear-end first.
3 Adaptations linked to feeding in insect eaters
3.1 Introduction
How evolution proceeds is obviously of central importance when studying mammals. Of fundamental importance to the way evolution works is the notion of natural selection, and in S182_3 Studying mammals: chisellers I'll be talking about what most researchers regard as this 'single most important idea in biology'. But before that, I want to describe some of the adaptations evident in insect eaters.
From your reading of LoM you'll appreciate that natural selection promotes the evolution of adaptations, which are heritable features of animals that increase the likelihood that they will survive and reproduce in the environment in which they live. Adaptations linked to obtaining food are of crucial importance and I'll be outlining examples from insect eaters in the following sections.
3.2 Shrews
Activity 2
Drawing on the notes you made about shrews in Activity 1, describe in a couple of sentences the water shrew's strategy for searching for and catching insect larvae.
Discussion
The water shrew dives into the water and swims rapidly, searching for food by using its long, sensitive whiskers to locate prey, such as insect larvae. Once located, an insect is grabbed with the teeth and taken to the surface to be eaten.
Efficient swimming is important for the water shrew because the time that an air-breathing animal can spend under water is limited. Water shrews spend only about 4-15 seconds under water during a dive and for their mode of feeding, speed is essential.
Let's think for a moment of the features of the water shrew that can be regarded as adaptations for swimming under water. First, oily fur prevents the coat from becoming waterlogged during a dive, and traps air bubbles. The water shrew's oversized feet with hairy ankles and digits act like flippers while the animal is submerged.
If a feature of an animal is an adaptation, then it should be possible to relate the structure of the feature to its function in the animal's life. The oil on the fur prevents wetting, an important function for an animal that dives into cold water. The air that is retained offers a degree of insulation. Insulation helps reduce loss of body heat while the animal is submerged and on land; cold ambient temperatures on land would further chill a small, wet animal emerging from the water.
Links between body structure (here, of feet) and function (here, for swimming) of this type are strong indicators that particular features are adaptations. Further evidence to support such a view comes from comparisons with similar animals that do not have these features. For example, the common shrew catches insect prey on land, and its feet are smaller in comparison to those of the water shrew, and it does not have hairy ankles.
3.3 Moles
In this section, you will meet one of the ways in which scientists are very precise about the way that they use ordinary words. Spot the difference between 'The golden mole evolved webbed hind feet so that it can shovel sand backwards as it moves forwards' and 'The golden mole evolved webbed hind feet and it uses them to shovel sand backwards as it moves forwards'. Why should such a subtle difference matter?
Activity 3
Watch the sequence on the golden mole from the TV programme (07.14-08.56) and jot down notes about the animal's main features, and how they link to searching for insect prey. In up to 100 words, describe which of the golden mole's features can be regarded as adaptations for burrowing in sand.
Discussion
The head is wedge-shaped, covered with thick leathery skin, and used to push through the sand. The eyes are tiny and covered with furry skin. Ear openings are covered with fur, protecting sense organs from sharp sand grains. The front legs end in flipper-like claws, used for swimming through the sand. The golden mole's hindlimbs are webbed, used for shovelling sand backwards as the animal moves forwards. (It seems appropriate that a swimmer should have webbed feet!)
The golden mole bears a strong resemblance to the marsupial mole, which lives in desert country in Australia. The marsupial mole 'swims' through sand in much the same way and feeds on insects and earthworms. The head is wedge-shaped and covered with a tough horny shield. The eyes and ears are very small. The front limbs end in two large flipper-like claws (shown clearly in LoM p. 25), which closely resemble those of the golden mole.
Like the golden mole, the marsupial mole has webbed hind feet, which are used to shovel away sand during 'swimming'. But marsupial and placental mammals are taxonomically very distinct. As I've mentioned in Section 1, the 'branch point' in the evolutionary tree - where these two groups separated from a common ancestor - was probably more than 90 million years ago.
These features of the golden mole and the marsupial mole are examples of convergent evolution, in which similar adaptations have evolved in two distinct and distantly related groups. Convergence is regarded as strong evidence that particular features are adaptations for a particular way of life. The similarities of the front feet of the two types of mole provide strong evidence of convergence, an impression reinforced by the fact that the forelimbs are not identical in every regard. Both animals have very short front limbs, with huge flattened claws, but in the marsupial mole, these are formed by fusion of the third and fourth digits, while in the golden mole they are the fused second and third digits.
3.4 Anteaters
Activity 4
View 'The Insect Hunters' on the DVD from 22.40-26.54, which shows the giant anteater, and make notes on what you see. On the basis of your notes, what features of the giant anteater could be regarded as adaptations for eating a diet composed of ants and termites? List each feature and state how the structure relates to its function (Figure 3 will be helpful to you). Your answer should be less than 100 words in total. (You will use your notes again in Activity 7.)
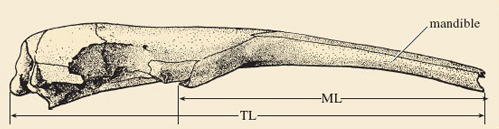
Discussion
The following features are adaptations for eating ants and termites.
Claws: Huge claws on the front feet are used to rip open termite hills and ant nests.
Jaws: The giant anteater has no teeth, even in adulthood; the jaws are fused and drawn out into a long tube supporting a long snout, which is inserted into holes dug out of termite mounds and ants' nests (Figure 3).
Tongue: The salivary glands are enlarged and secrete a sticky saliva that coats the long tongue, which is used to pick up ants and termites.
Comparison of the giant anteater with a similar-sized animal with no claws that cannot rip open termite mounds, e.g. a sheep, supports the view that anteaters' claws are an adaptation. But consider the following statement:
'The giant anteater had to evolve enlarged sharp claws so that it could rip open termite mounds.'
This statement implies that evolution occurs in a specified direction. However, evolution does not proceed with an aim or purpose, but primarily by means of natural selection. Statements implying a purpose for evolution are described as teleological. It is tempting to write about evolution in such a reassuringly 'commonsense' way and sometimes you'll come across statements of this type in LoM and the TV programmes. But on strict scientific grounds, it is not correct; it conveys an apparent belief that evolutionary change is forced in particular directions, perhaps by the 'needs of animals'. If, as a student of evolution, you use such language, others might conclude that you believe in 'directed evolution'. As you'll appreciate when we discuss in more detail how evolution is driven by natural selection operating on random variation, this concept is exactly contrary to how evolution comes about. So, the statement should be rewritten as:
'The giant anteater has large sharp claws on the front feet, used to rip open termite mounds.'
SAQ 5
Criticise and rewrite the following statement: 'The giant anteater evolved long tube-like jaws so that the snout could be inserted into holes dug into termite nests'.
Answer
The statement implies that evolution occurs with a purpose, and is teleological. It could be rewritten as: 'The giant anteater has long tube-like jaws supporting a long tubular snout that is inserted into holes dug into termite nests'.
3.5 Bats
There are two more activities in this section that give you more practice in writing. You will see that you are again given an approximate number of words to aim for in your answer. This number is a guide to the level of detail required - you will often find the same thing done in course assessment questions. Write your answer as concisely as you can, checking that you have not repeated any points, and then count the words. If you have written more than the suggested number, do not spend time going through your work again, cutting out the odd word here and there; that is not necessary at this stage, but do compare your answer with the one given, and try to identify where you could have cut down.
From LoM you'll know of the spectacular adaptations in bats (order Chiroptera) that relate to hunting and catching insects in flight. The major adaptations of bats that you read about are, of course, the development of wings and the structures linked to the production and detection of ultrasonic sounds. The bat wing is a modified mammalian forelimb, in which over evolutionary time, the bones have become considerably lighter and certain bones have undergone great elongation. It's revealing to compare a less highly modified forelimb from a human (Figure 4a) with that of a bat (Figure 4b). The bat's finger bones - metacarpals and phalanges - are long and thin. The ulna is greatly reduced, almost completely lost. The clawed thumb remains free and is used for moving around roosts. You'll notice that the bones within the human and bat forelimb are identically named. This is because they are homologous, i.e. they were derived from an equivalent structure in the ancient common ancestor of these two species. By contrast, the wings of bats and insects, though they share the same function, did not originate from the limb of a common ancestor; insect wings are extensions of the outer covering of the body, the cuticle. Thus insects and bat wings are termed analogous, i.e. identical function but without a common evolutionary origin.
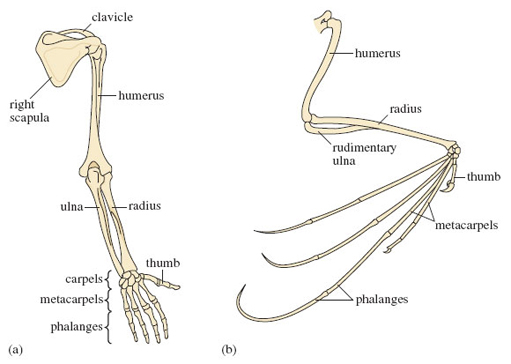
The concepts of homology and analogy were central to comparative biology from their invention early in the 19th century until very recently. These long established principles were challenged in the 1990s by the discovery that similar, in some cases identical, genes direct the development of eyes, legs, wings and other structures in animals as different as insects and mammals. The results of this very active field of current research are already prompting a fundamental revision of our thinking about homologous and analogous structures.
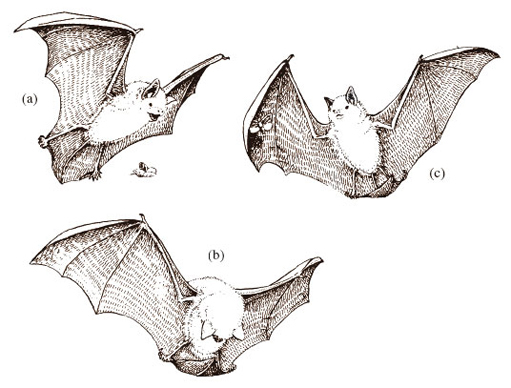
DA describes the bat's wing as an extension of the skin 'from the side of the neck across the fingers and down to the leg' [p. 45]. Some species have a tail membrane too, which extends between the two hindlimbs; Daubenton's bat (Figure 5) is one such species. The wing and tail membrane is a living tissue, comprising brown or black (and almost hairless) skin that has considerable elasticity. The wings are flexible, but are held relatively stiffly (i.e. under tension) during flight by the arrangement and action of those muscles of the bat devoted to flight.
Activity 5
Reread the part of LoM that explains how bats use ultrasonic sound as a navigational system and as a means of locating their insect prey during night flying [pp. 45-46]. Watch the sequence on bats from the TV programme, from 28.50-39.38, and jot down notes on how bats detect and catch insects. Explain, in up to 200 words in total, (a) how bats generate focused beams of ultrasonic sounds, and (b) how they use the sounds for navigation and locating prey (flying insects).
Discussion
(a) While flying, bats generate short bursts of ultrasonic sound with the larynx in their throat, emit them via the nose and mouth and listen to the echoes of the sounds as they bounce back from objects in the area. The beam of sounds is focused by means of a nose leaf in many bats. The shape of the nose leaf varies according to species, and includes bowls, slits and other shapes. The nose leaf may be mobile, enabling the bat to vary the width of the beam that is projected.
(b) The time taken for the echo to return to the bat varies with the distance from each object and whether it is moving or not. The type of echo received also gives a clue as to the nature of the object. So as the bat scans its surroundings during flight, it is able to identify flying insects. As the bat closes in on its prey, the 'short stabs' of ultrasonic calls become more frequent.
The human ear cannot detect these ultrasonic sounds, but researchers use bat detectors to track bats in flight. One type of detector picks up bat calls by an ultrasonic microphone and converts the sounds into sonic frequencies that are audible to the human ear. The output of a bat detector is, therefore, not an exact reproduction of the original bat call. Nevertheless, the patterns of sounds are reproduced, and are typical for each species, and also for the activity of the bat. Bat detectors have been used to look at feeding in Daubenton's bats, which featured in the TV programme. When these bats approach their prey, they emit intense bursts of sound, 'feeding buzzes', used for locating the insect's exact position.
Activity 6
Look back at the notes you made for Activity 5, above, and use them to write a brief description, in up to 150 words, of the prey-capture techniques used by bats featured in the TV programme from 28.50-39.38.
Discussion
Bats, of course, use the mouth, and in particular the teeth, for capturing prey but also, more surprisingly, the wings, which, as I've already mentioned, are very flexible. Variations of hunting strategies are seen in different species. Daubenton's bats fly close to the surface of a stream, grabbing flying insects with their mouth or feet and using the tail membrane as a catching basket (Figure 5b). The bat bends its head towards the scoop made by the tail membrane and picks up the insect with its teeth. Long-eared bats fly amongst branches of trees, and detect moths by listening for the sounds they make in flight or when eating; they use their teeth to pick up insects from a leaf, cupping the wings over the prey to prevent its escape.
4 Thinking about adaptation
Section 3 identified a range of adaptations in insect eaters, most linked with their mode of feeding. Particular structures are identified as having particular functions. But there are problems with the concept of adaptation if it's taken too far. Not all features of an organism have to be functional in ways that perfectly suit that organism to its environment. In fact, the whole notion of the 'perfection' of animals and their workings is best avoided. The TV programme shows, for example, some of the shortcomings of the hearing mechanisms that long-eared bats use to locate their prey.
Remember too that a particular species is 'here and now' - it's at one point in its evolution. Species very often retain structures that are vestiges of their evolutionary past. For humans, the appendix (part of the large intestine) is one such organ you might be familiar with. If you study evolution in greater detail, you'll come across many other examples of such 'vestigial structures,' as well as numerous examples where we have little idea about why and how a particular behaviour or complex structure arose and its role in a mammal's natural habitat. Being enthusiastic about the wonders of mammalian life, as LoM and the TV programmes very properly are, is fine, but don't run away with the impression that mammals are perfect, either in the sense that they are the most efficient and complex systems imaginable or that they are ideally suited to every aspect of their environment. This notion is so important that we'll return to the idea of adaptation in the next course in this series, when we will use further mammalian examples to look critically at the meaning of the term.
5 Temperature regulation and the consequence of size
5.1 Introduction
If you have already worked through S182_1 Studying mammals: a winning design, you'll be aware (from Section 5) that animals break down their food for conversion into usable forms of energy; thus, breakdown of food is sometimes called (as in the commentary to the TV programme) the 'internal fire'. Fire is a useful analogy because within the body, food is oxidised. This process is comparable to burning, but it is much, much slower and takes place in living tissue. Chemical energy released during breakdown of food is able to support growth, movement and processes essential for maintenance of life, such as excretion of wastes, beating of the heart and replacement of worn-out tissues such as skin and the lining of the gut. In endotherms (mammals and birds), the heat energy released during breakdown of food is used for maintenance of body temperature.
As defined in S182_1, an animal's metabolic rate is the rate at which it chemically breaks down food molecules and releases the energy they contain. For convenience, metabolic rate is often measured as the rate of uptake of oxygen per unit mass of an animal. The basal metabolic rate (BMR) is a measure of the rate of metabolism of an animal at rest, when the ambient temperature is such that no thermogenic response is evident.
You will have noticed from the TV programme that insect eaters have a huge range of body sizes. The smallest, a pygmy shrew, weighs about 2 g, in contrast to the giant anteater, which weighs up to 40 kg, and the giant armadillo, which can weigh up to 55 kg. Such differences in body size have profound implications for the regulation of body temperature and for food intake.
5.2 Body size and metabolic rate
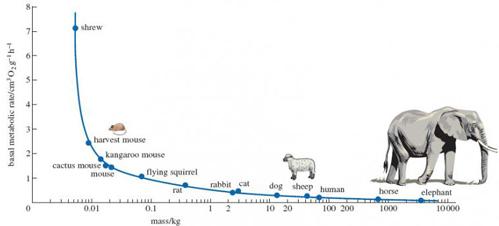
Figure 6 is a slightly more complex graph than those used in S182_1. In particular, the masses of the mammals that are plotted on the horizontal axis vary so much that a normal scale would squeeze together all the small and medium-sized ones, in order to be long enough to get the very large ones on the scale. So, as explained in the figure caption, a logarithmic scale is used here to get them all to fit. In this series of units, you won't come across plots like this very often, so it's not worth spending time struggling at this point to understand what a logarithmic scale means, if you are not familiar with one. You should still be able to read off the values of the plotted points as normal. As a general rule, you should be prepared to 'put to one side' complications like this that are not central to your learning and focus on the key points - here, the relationship between BMR and body mass. Don't forget to check the figure caption and look carefully at the units in which the measurements have been made.
It has long been known that the metabolic rate of mammals is closely related to body size. Look at Figure 6 which shows how BMR decreases in mammals as body size increases. (The way the units of body mass are marked onto the horizontal axis may well be unfamiliar to you, but you should find it possible to use the scale to work out the value of plotted points, most readily those between 1 and 10 and 10 and 100 kilograms.)
SAQ 6
From Figure 6 read off the BMR of the shrew, which weighs around 0.01 kg, and of the sheep, which weighs around 40 kg, and compare the values you obtain. Explain how your comparison relates to the energetic lifestyle of the shrew and what you know about the lifestyle of the sheep.
Answer
A shrew with a body mass of about 0.01 kg has a metabolic rate of about 7 cm3 O2 g−1 h−1. The BMR of a sheep is much lower, about 0.3 cm3 O2 g−1 h−1. You'll know of the very fast pace of a shrew's life from the TV programme, so its high metabolic rate may come as no surprise. High metabolic rates derive from rapid breakdown of food, which in turn requires relatively high food intake. Shrews spend nearly all of their time hunting vigorously for insects and eating them. In contrast, sheep have a slower lifestyle - long periods of rest interspersed with grazing - and have a lower BMR.
The daily food intake of the pygmy shrew reveals just how much a shrew eats; it consumes up to 1¼ times its body weight daily. Shrews need to feed every few hours, otherwise they die. A giant anteater weighing 40 kg would need to eat up to 50 kg of termites per day if it ate an equivalent amount to that taken in by the shrew. However, a giant anteater's average consumption is about 30 000 termites per day. Assuming a body weight per insect of about 0.03-0.05 g, 30 000 termites amounts to about 1-1.5 kg food in total. DA's figures suggest a much higher value for the giant African pangolin's daily intake of insects [p. 58] but even this amount comes nowhere near the shrew's intake, relative to body size.
5.3 Body size and surface area
You will be using some more maths in this section. Remember that areas are measured in units such as m2, which is read as metres squared or, more usually, square metres. Volumes are measured in units such as m3, which you should read as metres cubed or, more usually, cubic metres. Most measurements in science have units attached to them and you should never write down a number without its unit, if it has one. Some numbers don't have units, such as the ratio of one number to another. For example, if you had two jugs, one with a volume of 1500 cm3 and another with a volume of 500 cm3, the ratio of their sizes would simply be 3 (with no units), meaning that the first jug has three times the volume of the second one.
Why should the amount of surface that a mammal presents to its environment be of interest? Because body heat can be either lost to or gained from the external environment via the body surface. The larger the surface area of a mammal, the greater the potential rate of heat loss or gain. Let's concentrate for the moment on problems of heat loss. The fact that body temperature (i.e. Tb, a symbol introduced in S182_1) is so very often greater than outside, or ambient, temperature (Ta) means that frequently the direction of heat 'flow' is from the animal to the outside, i.e. heat is lost from the animal. (Other factors influence the direction of heat flow, such as how much solar heat is absorbed, but I'm ignoring them in order to keep things straightforward.) For a mammal struggling to regulate its body temperature at a relatively constant value, heat lost to the outside, via the surface, must be replaced by heat obtained from the breakdown of food.
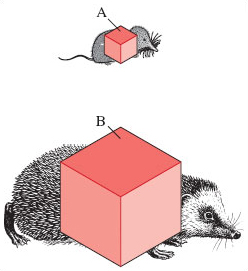
First, let's consider how the surface area of an animal relates to body size. We'll look at two mammals of very different body size; their relative proportions can very approximately be represented as two cubes of different dimensions, as in Figure 7. Follow the arithmetical steps carefully:
The smaller species, the shrew, is represented by cube A with a side of length of 4 mm.
The area of each cube face is 4 mm × 4 mm = 16 mm2.
As there are six faces to the cube, the total surface area is 16 mm2 × 6 = 96 mm2.
The volume of cube A is: 4 mm × 4 mm × 4 mm = 64 mm3.
The ratio of surface area to volume for species A is therefore 96/64 = 1.5. So for the shrew, the value for the surface area is 1.5 times greater than the value for the volume.
The larger-sized hedgehog is represented by cube B in which each side is 25 mm long.
This gives a total surface area of 25 mm × 25 mm × 6 = 3750 mm2.
The volume is of cube B 25 mm × 25 mm × 25 mm = 15 625 mm3.
The ratio of surface area to volume is 3750/15 625 = 0.24.
SAQ 7
What can you tentatively conclude from Figure 7 about the effect of body size on the ratio of surface area to volume and the resultant heat loss?
Answer
The smaller the animal, the larger is the surface area to volume ratio. Therefore, a small animal like a shrew has a greater potential for losing heat from its body because of its relatively large surface area.
In Figure 7, I've been a bit 'free and easy' with my estimates of body dimensions. The figure is a rough approximation - good enough for my purposes. I was tempted to draw in a third cube, representing a very sizeable insect eater such as the giant anteater. But even if I kept to only roughly the right proportions, the animal's outline and the corresponding cube would occupy a full page, so I hope the point is clear enough from Figure 7 as it is.
There's an important inference to be drawn here, which helps explain the relationship in Figure 6. A relatively large surface area (as with the shrew) will lead to a greater heat loss per unit mass of animal. In order to compensate for this enhanced heat loss, a mammal's metabolic rate has to be sufficiently high to maintain its body temperature at a steady value of 36 °C or so. Hence, generally speaking, the more massive an animal, the lower its heat loss relative to its size. In this sense, very large animals have relatively low running costs. Their food requirements are modest - not in the absolute sense of the total amount of food ingested, measured say in kilograms, but relative to their body size, i.e. kilogram of food per kilogram of body mass.
One way of avoiding excessive heat loss is to increase insulation. Rather than constantly eating, why don't small mammals increase insulation? The coat of small mammals simply cannot be thick enough to provide adequate insulation in cold conditions. Thickness of the fur coat is limited by the amount of fur animals can physically carry. If a shrew had an effective winter fur coat, its feet would not reach the ground! For large insect eaters living in temperate climates, e.g. anteaters, effective insulation by fur is more feasible.
6 Strategies for coping with cold and food shortage
6.1 Toughing it out
For small insectivorous mammals, the best solution to the problem of obtaining sufficient energy for their requirements, notably the maintenance of body temperature, is to feed continuously. But in temperate regions, insect eaters have to cope with shortage of insects during cold winters. In Section 6 I want to focus on two strategies that insect eaters use to cope with problems of overwintering.
When talking about the giant anteater, the TV programme commentary claims that these animals are short of food all the time, because their diet of termites is not very nutritious.
Activity 7
Look back at the notes you made on the giant anteater sequence in Activity 4 (TV programme 22.40-26.54 ) and explain how the giant anteater reduces its energy expenditure.
Answer
Giant anteaters spend much of their time sleeping ('asleep for 15 out of 24 hours'), during which they cut down loss of heat by covering their body with their long bushy tail. Giant anteaters maintain a lower body temperature (just 32 °C) than most other mammals, so they do not need to use so much of the energy released from food breakdown for maintaining body temperature.
Shrews do not have a coat of long fur and they have a high metabolic rate (Figure 6), maintaining a Tb of 37 °C. The insects that shrews eat (mostly found in the leaf litter) are often not hugely depleted during the winter but, as you'd expect, shrews are less active over this period, compared to the summer bonanza. Indeed, shrews may undergo brief periods of torpor in winter. Torpor is a state of lethargy during which the animal allows its body temperature to drop.
Torpor lasts for short periods only, either at night or during the day. As the torpid animal is not using energy to maintain its normal high level of activity or high body temperature, significant amounts of energy are saved. Savi's pygmy shrew becomes torpid at low temperatures. Researchers measured a Tb of just 12 °C for Savi's pygmy shrews during torpor at Ta of 4 °C, compared to Tb of 36 °C during normal activity. Heart rate was just 100 beats per minute during torpor, compared to 800-1200 per minute during normal activity. A more profound adjustment to winter shortages would be to enter true hibernation, when Tb would fall dramatically. And yet no shrew hibernates - why not, when 'opting' out would be presumably so advantageous?
6.2 Opting out
This last section of the course contains, I think, some of the most challenging science that you have met so far. Take it slowly, translating all the abbreviations in your head as you come to them (read BAT as 'brown adipose tissue', for example) and looking carefully at the graph in Figure 8 and at Figure 9, remembering to read the captions. Don't allow yourself to get 'bogged down' if you don't understand something first time; continue to the end of the section and then read the advice in the box at the end.
Hibernation is a more radical and extended adjustment than torpor. Insect eaters that are able to store sufficient amounts of fat in their bodies can undergo hibernation in response to the low ambient temperatures of winter. For shrews, a combination of high metabolic rate and small body size means that this particular physiological feat seems beyond them. By contrast, although some bats display torpor on almost a daily basis, many other species, especially those living in cool climates, undergo genuine hibernation.
Activity 8
Watch 'The Insect Hunters' from 39.58-42.54, which shows overwintering in little brown bats, and make brief notes. From your close observations, especially of the pictures from the thermal imaging camera, describe how the bats hibernate and then define hibernation.
Discussion
During hibernation, the bats are very cold and hang upside down in a profound form of sleep. Periodically the bats warm up and arouse, in order to drink water - though the males might indulge themselves in other respects. On this evidence, hibernation is a prolonged winter sleep in a sheltered but cold spot, during which the animal's body temperature plummets, interrupted by brief periods of arousal. Thus, true 'deep' hibernation consists of successive bouts of torpor, interrupted by arousal.
The flying insects preyed upon by bats are scarce or absent in winter, the very time when bats would need extra energy to maintain a high body temperature. Before hibernation, bats build up stores of fat in their body, and some such stores have an important role during arousal, as I'll explain shortly.
Biologists used to believe that hibernating animals simply allowed their body temperatures to drop passively to that of the burrow or cave in which they spend the winter. More recent research has shown that body temperature is tightly controlled during entry into hibernation and during hibernation itself. Data from a much larger insect eater, the hedgehog, is of special interest.
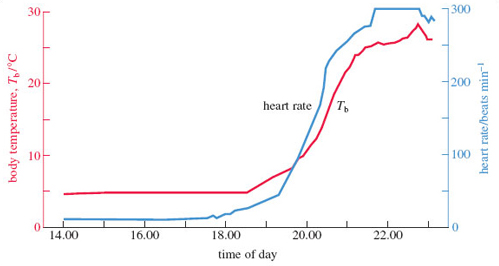
Hedgehogs in Britain hibernate from around November to April, with precise timings dependent on weather conditions. They become aroused from hibernation about every 7-11 days. Temperature and heart rate during hedgehog hibernation have been measured for long periods via miniature sensors implanted into the animal. Figure 8 shows body temperature (Tb) and heart rate readings during a period of deep hibernation (for four hours from 14.00 to 18.00, i.e. 2 p.m. to 6 p.m.), and during a subsequent period of spontaneous arousal (18.00-23.00, i.e. 6-11 p.m.).
SAQ 8
Read off the graph the values of Tb and heart rate before arousal. (Figure 8 has two different vertical axes; the left axis shows the values for Tb, so run your eyes to the left as you read off the Tb values. Refer to the right axis for heart rate values.)
Answer
During deep hibernation, Tb of the hedgehog was about 4 °C. Heart rate is more difficult to determine but is a very approximately 10 beats per minute.
Compare the very low heart rate during hibernation with the normal heart rate of 250-300 beats per minute when the hedgehog is active. Breathing rate is also reduced sharply in the hibernating hedgehog, down to just one breath per hour, compared to 25-50 breaths per minute in an active individual. Prolonged periods of breath-holding, known as apnoea, have been observed in hibernating hedgehogs, with a 'record' breath-hold of 150 minutes.
A hibernating hedgehog may appear lifeless and not subject to any physiological control, but this is far from the case. When not hibernating, the hedgehog, like all mammals to a greater or lesser extent, maintains its Tb around a set-point. If Tb falls below or rises above 35-36 °C, physiological mechanisms come into play to warm or cool the hedgehog's body as appropriate. You can think of the control mechanisms as being similar to those that control a central heating system to maintain your house at a comfortable temperature. If the temperature of a room falls below that set by the thermostat, the heating is switched on; if the room temperature is too hot, the thermostat switches off the heating. The thermostat of the hedgehog has no known physical basis but it is clear that heating and cooling of the body is controlled by a special area in the brain, the hypothalamus. If a non-hibernating hedgehog gets too cold (Tb dropping below 35-36 °C), the heating is switched on - shivering and increases in metabolic rate (see course S182_1) take place. If the body temperature of the hedgehog exceeds the set Tb, the heating mechanisms are switched off.
During hibernation, the set Tb is reduced to just 4 °C, analogous to turning down the thermostat to 4 °C in your central heating system. If the hedgehog's Tb drops below 4 °C during hibernation, the animal will react by increasing its heart and breathing rate and its rate of metabolism, generating sufficient extra heat to raise its body temperature. Arousals are provoked by chilling but, as you have seen, many arousals occur spontaneously.
SAQ 9
Figure 8 shows changes in heart rate and Tb during spontaneous arousal, beginning at 18.00. Describe the pattern of changes shown in the graph.
Answer
Heart rate increases from below 10 beats per minute to a maximum of about 300 beats per minute by 23.00. Tb increases from 4 °C to a maximum of over 25 °C at 23.00.
Why hedgehogs, and hibernators in general, become aroused periodically is not fully understood. Hibernating hedgehogs spend a total of about 80% of the time with a very reduced Tb, waking up between 15-22 times during a hibernation period.
During arousal, the body is warmed up by fat breakdown in BAT (brown adipose tissue or brown fat), which is very different from the more familiar white fat (white adipose tissue, or WAT). What we commonly think of as fat consists of masses of WAT cells, each loaded with a large round fat vacuole (Figure 9a). Traditionally, WAT has been viewed as a storage tissue that animals draw on during prolonged exercise, or in 'emergencies' when they are short of food. (Recently, WAT has been found to have more complex structures and a wider range of functions than was believed.) Breakdown of the fat contained in WAT into its constituent building blocks (the most significant types are called fatty acids) is the first step to releasing the chemical energy used to fuel metabolism. Prior to hibernation, hedgehogs and bats build up stores of WAT. BAT normally builds up too during this period.
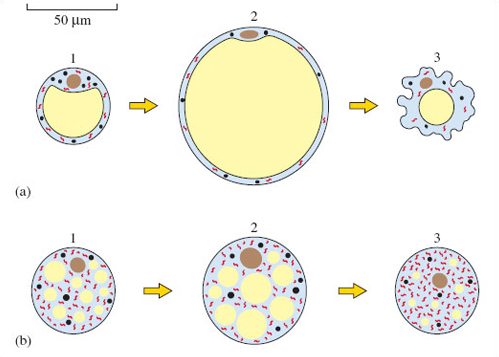
BAT consists of masses of somewhat smaller cells, but each has a number of small fat droplets, and also reserves of a 'storage' carbohydrate, glycogen (Figure 9b). What is remarkable about BAT is that its primary function is the generation of heat. In most other types of cell, metabolism is geared in such a way that the breakdown of fuels such as fatty acids and glucose releases energy in a chemically useful form. This 'captured' energy can be used within such cells to maintain all the energy-demanding 'housekeeping' activities, such as synthesising macromolecules, or transporting different chemical building blocks around the cell. But the breakdown of fatty acids in BAT (the final stages of which occur in the mitochondria) releases energy not so much in the form of these neat 'parcels' of chemical energy but as heat. Once BAT becomes switched on in this way, blood circulating through BAT is warmed and the warmed blood circulates throughout the body, raising Tb. Indeed, the endearingly termed 'bat BAT' has a vital role in arousal.
SAQ 10
From your observations on thermal imaging of the little brown bats during arousal (the TV programme extract from 41.40-42.52), suggest the location of BAT in these animals and describe the spread of heat throughout the bat's body.
Answer
Thermal imaging shows a large hotspot behind the neck, between the shoulder blades, the likely position of a large depot of BAT. Heat released by breakdown of fat (and glycogen) in the BAT spreads around the body. The head appears to be the warmest; the rest of the body and the limbs less so.
So the use of thermal imaging provides a wonderful insight into the role of BAT in arousal of hibernating bats. But why arousal is necessary remains a mystery, especially since it uses up so much of the precious energy reserves of the body. Little brown bats do not feed during arousal, but as you've seen, they drink water. Perhaps doing so helps them get rid of accumulated toxic wastes via urination, but this explanation is by no means certain. This question is just one further example of just how little is known about bats, which only increases a sense of wonder about these intriguing mammals.
How did you get on with this last section? If you found it difficult, don't worry. Read through it once more and see if that helps. If you plan to study the next course in this series, move on to S182_3 Studying mammals: chisellers and promise yourself that you will come back and reread the section later. You will probably find that it seems much more straightforward when you look at it again.
Now that you are at the end of this free course, read through the learning outcomes in the course introduction and consider making notes on them
At this point, you may feel that you would like to explore some aspects of insect eaters in more detail. The BBC website provides links to valuable resources for obtaining much more information about mammals of all sorts and if you are able to, now might be a good time to investigate what is available.
Conclusion
This free course provided an introduction to studying Environment & Development. It took you through a series of exercises designed to develop your approach to study and learning at a distance, and helped to improve your confidence as an independent learner.
Acknowledgements
Course image: Paislie Hadley in Flickr made available under Creative Commons Attribution-NonCommercial-ShareAlike 2.0 Licence.
Grateful acknowledgement is made to the following sources for permission to reproduce material in this course:
The content acknowledged below is Proprietary and is used under licence.
Figure 2a adapted from Reeve, N. (1994) Hedgehogs, illustrated by Ruth Lindsay, T. & A.D. Poyser Natural History Series;
Figure 3 adapted from Young, J. Z. (1962) The Life of Vertebrates, Oxford University Press;
Figure 5 adapted from Richardson, P. (1985) Bats, Whittet Books;
Figure 6 Adapted from Schmidt-Nielson (1975), Animal Physiology, Adaptation and Environment, Cambridge University Press and reprinted in Randall et al. (1997) Eckert Animal Physiology: Mechanisms and Adaptations, W. H. Freeman and Company;
Don't miss out:
If reading this text has inspired you to learn more, you may be interested in joining the millions of people who discover our free learning resources and qualifications by visiting The Open University - www.open.edu/ openlearn/ free-courses