

3.1 Signals for entry
Despite the fact that hibernation is reflected in a number of profound and operationally distinct physiological changes, changes in T b continue to be the recognized signs of its onset, interruption and termination, because of the relative ease of monitoring T b. Onset is triggered both by endogenous and exogenous cues.
Question 6
What would you identify as exogenous cues?
Answer
The three most important environmental stimuli initiating torpor are food supply, daylength and T a, though the order of their importance differs between species and between seasonal hibernation and daily torpor.
Both food supply and the amount of body fat are relevant. In the short term, a diminution of available food towards the beginning of the hibernation period may itself trigger hibernation, and in a laboratory cold room at constant temperatures, torpor can be induced in several species by the removal of food. Examples here include some hummingbirds and the poor will, where entry into torpor rapidly follows the removal of food. In the longer term, food supply determines the animal's ability to build fat reserves, and in some species, including some hamsters and ground squirrels, the presence of large fat reserves may be necessary for the animal's entry into hibernation.
However, others may not readily enter torpor in the absence of a store of food. Siberian hamsters (Phodopus sungorus) may fail to enter torpor even when injected with doses of insulin that result in a large and long-lasting fall in blood glucose. In this species in the wild, as well as others such as chipmunks, there is evidence that daylength is of greater importance as a cue. In general (and perhaps always), decreasing daylength is part of the stimulus, but the mechanism by which it acts may involve several separate routes. The first, which is not yet well understood, is that decreasing daylength causes an increase in the secretion of the peptide hormone melatonin from the pineal gland in the brain. Melatonin appears to act via a number of routes to predispose the animal to torpor. The second route is via the hypothalamus and the gonads; in most hibernators, entry into hibernation does not take place if there are high levels of androgens in the blood, and in the Turkish hamster withdrawal of the testes into the body cavity is a prerequisite for hibernation in males. Likewise, an injection of androgen into a torpid male hamster provokes arousal. It may also be that melatonin from the pineal gland acts to reduce gonadal secretions, as well as acting directly on the brain. Once again, the importance of ambient temperature as a cue to entry into hibernation may vary between species, though no species enters a bout of torpor unless the T a is below its thermoneutral level.
Animals such as hamsters and chipmunks are sometimes called facultative hibernators because they hibernate in response to environmental conditions. This contrasts with the so-called obligative hibernators, such as the ground squirrels and marmots, whose sequence of fattening, hibernating and arousing seems to be strongly driven by an endogenous annual cycle under physiological control. Figure 10 is drawn from data on the golden-mantled ground squirrel (Spermophilus lateralis; Figure 5) (Strumwasser, 1960), kept in the laboratory for 2 years under constant conditions of light (12 hours light and 12 hours dark) and at a constant temperature (22° C).
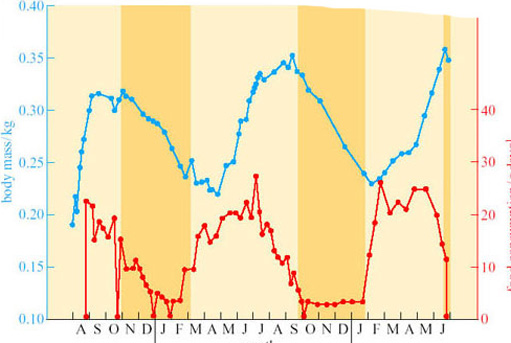
Question 7
What several conclusions on the triggers for hibernation can you draw from this figure?
Answer
As the sequence of food consumption, weight gain and hibernation continue as normal under constant environmental conditions, environmental triggers are not essential for hibernation in this animal. However, it does appear that the cycle may be shortening: in the first year the animal entered hibernation in late October, in the second in late September, and at the end of the experiment it was just entering hibernation at the end of June. Thus, this figure suggests that although the timing of the cycle may be primarily due to an endogenous circannual rhythm, in natural circumstances the timing may be re-set annually by environmental factors.
Thus, a sharp distinction between environmental and endogenous cues to entry cannot be drawn: indeed, the importance of the state of gonadal activity to hibernation in the hamster already reveals the interrelationship of these cues.
Many hibernators show a marked cycle in the production of thyroid hormones, with a decrease in their secretion at times when a non-hibernator would be increasing secretion to increase thermogenesis. Further hormonal and neural controls over the hibernation cycle are described in Section 6.