

3.5 Arousal (continued)
Question 9
What alternatives to shivering might act as a source of heat?
Answer
BMR is maintained mainly by a number of tissues with high metabolic activity. One of these, BAT, is unique in its ability to adjust its output of heat from being very low to being the body's principal source (see Box 1).
Box 1: Background to brown adipose tissue (BAT) – the role of uncoupling protein
Experiments using rabbits show that a marked lowering of T a initially lowers the temperature in many parts of its body, including areas that contain BAT. However, the temperature of the area with BAT rapidly returns to normal (37° C) because of thermogenesis within the BAT. In contrast, muscle temperature continues to fall because it has no capacity for temperature-related thermogenesis. The temperature of organs surrounded by, or near to BAT, show a smaller decrease in temperature. These results explain how core temperature, and hence the functioning of essential organs, is protected.
Mammals contain white adipose tissue (WAT) and, usually, BAT. The proportion of the total mass that is BAT varies from species to species, some having none but some having up to about 5% of body mass. The extent to which BAT produces heat in response to cold stress depends on the amount of BAT. Broadly speaking, mammals with small neonates, small mammals that are cold-acclimatized, and mammals arousing from hibernation, all contain a significant amount of BAT and are substantially dependent on it for thermogenesis. BAT is very much more thermogenic than any other tissue, by about a factor of ten (per unit mass), and is located in discrete depots with a good vascular supply. These depots contain brown adipocytes, which have a characteristic size and structure and are well endowed with numerous, large mitochondria with many cristae and a high concentration of cytochromes. It is the degree of vascularization and the concentration of cytochromes that give BAT its brown colour. Brown adipocytes also contain more glycogen than white adipocytes. White adipocytes are mainly unilocular (one large vacuole containing fat), whereas brown adipocytes that are thermogenically active are multilocular (a large number of small vacuoles containing fat). Brown adipocytes uniquely contain uncoupling protein (UCP). BAT is well innervated by neurons of the sympathetic branch of the autonomic nervous system. Neurons innervating brown adipocytes contain the neurotransmitter, noradrenalin; those innervating blood vessels contain another neurotransmitter, neuropeptide Y.
WAT is primarily a store of fat that is mobilized to provide lipid fuel for tissues remote from itself. In contrast, the function of BAT during cold stress is that of thermogenesis. When a BAT depot is cold-stressed, there is rapid cell division that increases the mass of BAT. The diameter of blood capillaries increases, and there are also changes in the histological make-up. Cold stress also leads to an increase in BAT sympathetic nervous activity and in the tissue concentration of UCP.
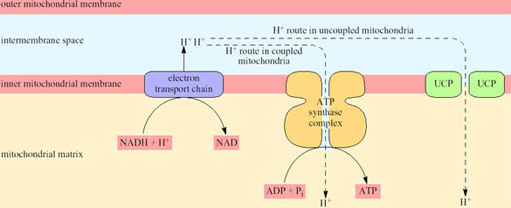
Non-BAT mitochondria (whether in WAT or any other tissue) are capable of transforming about 40% of the chemical energy of the respired substrates to the chemical energy of ATP; the other 60% appears as heat. However, because the rate of respiration is coupled to ATP production, the rate of respiration, and thus oxygen consumption and heat production, is comparatively low. ATP is produced by a mechanism that depends on the inner mitochondrial membrane being impermeable to hydrogen ions except via channels that are part of the ATP synthase complex. In BAT mitochondria however, fuel oxidation is uncoupled from ATP production, thus ensuring that all the energy released from oxidation is released as heat. As a consequence of uncoupling, which is brought about by the uncoupling protein UCP, the rate of heat production is greatly increased (Figure 20).
UCP is situated in the inner mitochondrial membrane and acts as a proton translocator providing a route by which hydrogen ions (built up in the intermembrane space as a consequence of electron transport) are able to re-enter the matrix without passing through the ATP synthase complex.
Blood flow to BAT reaches a maximum level in the arousal process, and recent work has shown that, while the uncoupling protein (UCP) in the mitochondria is in a ‘masked’ or inactive form during hibernation, the amount of active UCP is rapidly increased during arousal (see Section 4).
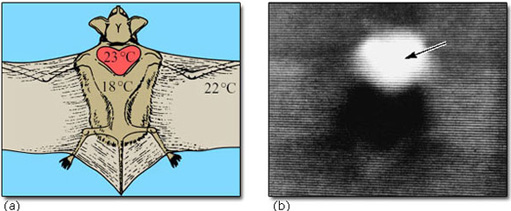
The interscapular region contains the largest mass of BAT near to the body surface. In hibernating mammals, this region is significantly warmer than other parts of the body (except possibly the heart) in the early stages of arousal. In big brown bats, the area of BAT is always warmer than the heart, and arousal is very fast (8° C to 37° C in less than 30 minutes). Figure 21 shows the thermographic tracing of body heat (infrared radiation) in an arousing bat. The area of BAT is the warmest part which suggests that it is the thermogenic source (Hayward and Lyman, 1967).
We should not infer that BAT is the major or only source of heat in all arousing hibernators. In those rodents which lack BAT, and probably also in the hedgehog, most of the heat in a normal arousal is generated by the shivering of skeletal muscle. Although birds do not seem to possess BAT, they are capable of non-shivering thermogenesis (NST), which is cold-induced heat production that is not due to muscle shivering. This kind of thermogenesis takes place in the muscles of cold-adapted ducklings and emperor penguin chicks, and there is evidence that it depends on free fatty acids liberated from WAT. Molecular evidence reviewed in Section 4 is pointing to the possibility that WAT, as well as BAT, can serve as a source of heat. However, it is not yet known what the relative importance of shivering and non-shivering thermogenesis is during arousal from daily (or more prolonged) torpor. Most birds can increase their BMR by a factor of four or five under extreme cold stress.