

5 Physiological adaptations – respiration and energy provision
5.1 Introduction
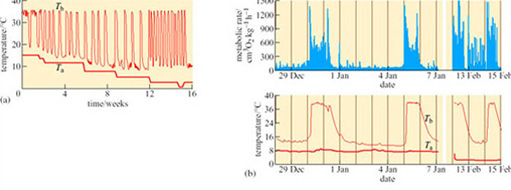
The change in BMR observed in all hibernators has traditionally been viewed as a passive response that is a consequence of hypothermia. However, many studies have provided evidence for temperature-independent regulation of BMR. In the alpine marmot (Marmota marmota), a BMR that is less than 5% of summer levels is maintained despite the frequent fluctuations in body temperature between 8 and 18° C. The mechanism of body temperature regulation in marmots, during long periods of hibernation, has become clearer following investigations of T b and BMR throughout this phase. Entry into hibernation is facilitated by a precipitate drop in BMR that precedes slower temperature changes, then throughout the winter, bursts of thermogenesis occur quite independently of T a (Figure 27) (Ortmann and Heldmaier, 2000).
It is widely acknowledged that mammals switch to the use of lipid from WAT during hibernation. A period of ‘fattening-up’ precedes the onset of hibernation, under the control of hormones which stimulate lipid storage. In mice induced to enter a near-torpid state, levels of leptin are low, reducing lipolysis and promoting lipid storage. However, BMR is lowered in the little brown bat (Myotis lucifugus; Figure 9), despite an increase in plasma leptin during the pre-hibernation period which suggests that weight gain is controlled by other hormones together with a resistance to leptin-induced satiety in this species.