

3.2 Penguins
Penguins (order Sphenisciformes) are an ancient and distinctive group of flightless, short-legged birds that evolved in the Southern Hemisphere, probably around New Zealand, about 65 Ma ago in the late Cretaceous, although the oldest known fossils date from about 45 Ma ago.
At a maximum body mass of more than 40 kg, the emperor penguin (Aptenodytes forsteri;Figure 10a) is the largest living penguin (some fossil species were much bigger) and is found further south than any other vertebrate. Like other penguins, emperors feed on fish, squid and large crustaceans that they catch by diving and chasing the prey underwater. The main predator of adult penguins is the leopard seals (Hydrurga leptonyx;Figure 10b), the largest and most agile antarctic seal, that has a varied diet including other seabirds and smaller seals, as well as fish, squid and crustaceans.
Emperor penguins breed on the iceshelf, away from predators such as skuas that take eggs and chicks, on breeding grounds that may be as far as several hundred kilometres from the open water. The males leave the feeding areas in early April (autumn in Antarctica) and fast during 6 weeks of courtship and for a further 2 months while brooding. Only one egg is laid, and the male carries it on his feet and broods it in a special flap of feathered skin that extends from his abdomen. Brooding penguins are inactive, keeping close together in large groups and walking an average of only 30 metres per day, thereby minimizing energy expenditure to near BMR. If his mate has not returned by the time the chick hatches, the male feeds his offspring on ‘curds’ formed from deciduous tissue in the oesophagus and broods it as he did the egg (Figure 10a). As soon as he is relieved by his mate, he walks back to the open water in what is by then midwinter, continuously dark and very cold.

The female also fasts during courtship, but she returns to the sea after presenting her mate with a single egg that is large relative to her own size. The female fattens quickly while at sea, eating 6–8 kg per day and increasing her body mass by about one-third, before returning to the breeding grounds to take her turn to feed the chick on curds and partially digested food regurgitated from her stomach.
René Groscolas and other French biologists from Strasbourg spent many months in Antarctica studying the physiological mechanisms behind these habits (Groscolas, 1982, 1986). Figure 11 shows the measurements that they made on wild penguins during the breeding season and in the following 3 weeks, while the birds were artificially prevented from returning to the sea to feed at the end of the natural fasting period. Every few days, marked penguins were caught, weighed, their rectal temperature measured, and a sample of venous blood taken.
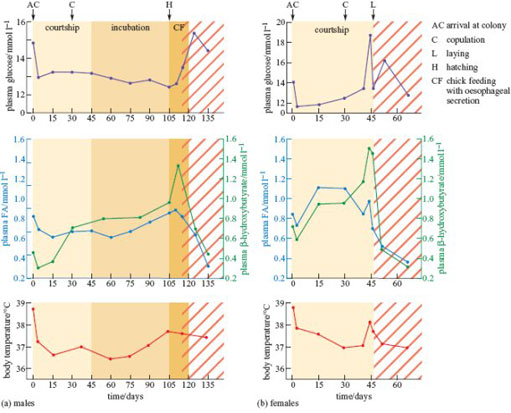
While fasting at the breeding colony, the mean body mass of the males fell by 40.5%, from 38.2kg to 22.75 kg, at an average rate of 35 g per day. After falling slightly during the first few days, the body temperature, and levels of glucose and fatty acids in the blood plasma were constant (Figure 11a), and well within the ranges of values measured in penguins that were feeding regularly. The ketone β-hydroxybutyrate is produced by partial oxidation of fatty acids and can substitute for glucose in some energy-producing pathways in some tissues. Its concentration increased steadily, reaching a peak when the fathers began to feed their chicks. The smaller females lost only about 22% of their initial body mass during their shorter fast. Except during the period of egg-laying, the pattern of changes is similar to that of the males.
SAQ 6
Why should egg-laying affect body temperature and metabolism?
Answer
Egg production involves the synthesis of large quantities of protein and lipid (for yolk), and the withdrawal of calcium stores (for shell formation), which generate heat and require levels of circulating glucose similar to those that support strenuous activity.
The tenfold increase in the concentrations of β-hydroxybutyrate is small compared to changes of up to 40-fold observed in the blood of pigeons, poultry and humans after just a few days of starvation. When artificially prevented from returning to the sea at the end of their normal fast, plasma fatty acid and β-hydroxybutyrate concentrations decreased sharply in penguins of both sexes. Their rate of weight loss also increased abruptly, reaching a mean of 542 g per day for the lightest penguins that weighed only 17.5 kg.
SAQ 7
What can you deduce from these observations about the penguins' fat stores and energy metabolism?
Answer
Production and utilization of free fatty acids decrease, probably because stores of triacylglycerols are almost exhausted. So the penguins start utilizing protein at a much higher rate. Because much less energy is produced from the breakdown of each gram of protein, a much higher rate of weight loss is necessary to meet the energy requirements of maintaining an almost constant body temperature.
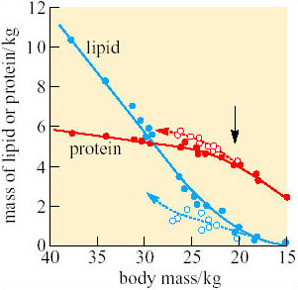
This conclusion is confirmed by increased excretion of uric acid during enforced starvation. Other measurements indicate that during natural fasting, 93% of the penguins' energy comes from oxidation of fatty acids released from adipose tissue (Figure 12). The small quantity of glucose needed to support glucose-dependent tissues (e.g. the brain) is formed mainly from the glycerol in triacylglycerols, and only small quantities of protein are utilized.
SAQ 8
Are the reserves replenished in the same way as they are depleted?
Answer
No. As shown on Figure 12, protein is withdrawn last during fasting but replenished more rapidly than lipid when the penguins start feeding again.
These observations suggest that, as in other animals, loss of protein has serious disadvantages and is only a ‘last resort’ used when other energy reserves are exhausted.
The mean body mass of male penguins leaving the colony is around 23 kg, which, from calculations based upon the data in Figures 11 and 12, indicates triacylglycerol reserves of about 2 kg. This amount is just sufficient to sustain the penguin as it walks, using energy at 2.8–4.5 times BMR, as far as 100 km back to the open sea. Under normal circumstances, the birds begin feeding just before exhausting completely the lipid in their adipose tissue (Figure 12). Utilization of protein reserves involves drastic alterations in metabolism and they do not last long, so if the weather is unusually severe, or the sea-ice is exceptionally extensive, or stocks of fish at the feeding grounds are low, penguins that were even slightly underweight at the start of the breeding season may not survive. Indeed, Groscolas suggested that the decrease in the plasma concentration of fatty acids and/or of β-hydroxybutyrate may be the metabolic signal (the black arrow on Figure 12) that prompts the parent to abandon its chick and return to the sea, even if its mate has not yet come back. Each year around 30% of eggs and chicks are abandoned for various reasons, and without parental care they always die. However, mortality among adult penguins is quite low, and each bird may breed many times during a long lifetime.
Comparison between different species of penguin shows that, in general, larger species can fast for longer, suggesting that the very large extinct penguins may have undergone fasts lasting many months.