

3.3 Integration of anatomical features and biochemical and physiological strategies in evaporators
Birds and larger desert mammals that use evaporative cooling risk dehydration because of the difficulty of finding sufficient drinking water. For mammals, evaporative heat loss includes panting and sweating.
In small mammals and birds the temperature of exhaled air is often lower than T b, resulting in condensation of water on the nasal mucosa. Small desert mammals rely on this mechanism for water conservation, while resting in their cool burrows during the heat of the day. However, for mammals and birds exposed to high T a, the nasal counter-current heat exchanger minimises water loss, and so works against the need to increase heat loss by evaporation of water (Figure 32).
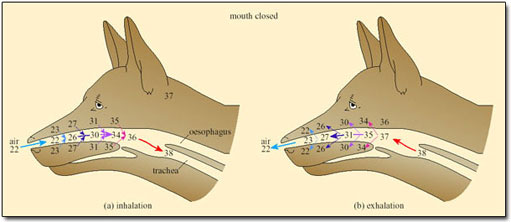
Only when T a approaches T b and the temperature of the inspired air reaches that of the body core, is the heat exchanger abolished. Even then, and for dry air, only about 12 per cent of metabolic heat is dissipated at T b of 38°C. As humidity increases, the proportion of metabolic heat dissipated declines.
Increasing the rate of ventilation of the nasal mucosa increases the rate of evaporation, but risks over-ventilating the lungs and blowing out too much carbon dioxide.
In dogs, foxes and other species that pant, evaporative cooling is promoted by opening the mouth; a simple mechanical device further increases the effectiveness of cooling by respiratory water loss. A valve at the back of the throat, driven by breathing movements, directs a large proportion of the air that was inhaled through the nose out through the mouth, thereby bypassing the nasal heat exchanger (Figure 33).
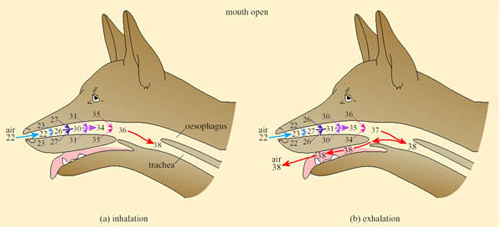
This mechanism can be used to modulate the rate of evaporative heat loss without affecting the respiratory frequency or volume and is exploited to the full during thermal panting under most heat loads.
During severe heat stress, breathing changes to a slower deeper second-phase panting, in which air passes out through nose and mouth. The dog's tongue is richly vascularised and the rate of blood flow increases with a rise in body temperature. Exposure of the buccal area means that there is no significant heat exchange in the mouth (unlike that shown in Figure 32) so the rate of evaporation is maximised during second-stage panting. Second-stage panting normally occurs in the dog during exercise, when both enhanced cooling and increased gaseous exchange in the lungs are required.
Panting is an important cooling mechanism for foxes and dogs that chase prey. The fennec fox (Fenecus zerda), a species found in the Sahara desert, is reputed to pant at 690 times per minute after chasing prey. Kit foxes reduce the need for panting by staying in dens during the day and hunting at night, or at dawn and dusk (Section 2.4). Rüppell's foxes (Vulpes rueppelli; Figure 34) live in the Rub' al-Khali of Arabia, the largest existing sand sea, which is an extremely arid desert with no permanent sources of drinking water.

Rüppell's foxes do not drink, but obtain all their pre-formed water from their food, supplemented by metabolic water production. By avoiding the need for panting during the day, Rüppell's foxes might be expected to have a reduced total evaporative water loss (TEWL) in comparison to fox species living in mesic habitats. Resting in a den during the day would reduce TEWL, but Rüppell's foxes would have to travel long distances at night while hunting prey, mainly rodents, birds and arthropods which would increase the need for evaporative cooling. Williams et al. (2002) measured TEWL of individual foxes in a specially designed metabolic chamber. Field water flux (the water flux under natural conditions in the field) was determined in individual foxes using the doubly-labelled water technique. The mean whole body TEWL for six foxes at 35°C at their basal metabolic rate was measured as 50.1 g water day−1. This value is about 55 per cent lower than expected from comparisons with other mammalian species of similar body mass. The researchers suggest that Rüppell's foxes are particularly efficient at reducing either cutaneous or respiratory water loss. Mean field water flux (FWF) per day in the six foxes was 123 ml day−1 with 26.1 ml water day−1 provided from catabolism.
Comparison of mass-adjusted values for water flux in Rüppell's foxes with values obtained for swift foxes living in grass prairie where water is more readily available, showed that water flux is 34 per cent greater in this mesic species. However, care is needed when making such comparisons between just two species, as we shall see in Section 4. The water flux of Rüppell's foxes is about 30 per cent less than that predicted by physiologists for a desert mammalian carnivore; the prediction assumed that desert carnivores would have higher rates of water flux than mesic species because of their higher rates of TEWL. It is tempting to suggest that the nocturnal habits of Rüppell's fox with consequent reduction in TEWL account for the low TEWL in this species.
Because birds of all sizes tolerate hot arid conditions, physiologists considered that desert birds, being diurnal animals exposed to extremes of ambient temperature and aridity in deserts, are successful because of their avian physiology, not because of specific adaptations. For example, as the normal range of core body temperature in birds (41–42°C) is higher than that in mammals, the need for evaporative cooling may not be as great as that in mammals. Because birds are uricotelic, that is, they excrete uric acid rather than urea, relatively little water is required for the excretion of nitrogenous waste. Uric acid is excreted as a paste, with a very low water content. It is relevant to note that carnivores, which have a high protein diet, produce relatively large quantities of urea as a waste product. This urea increases the osmolarity in the kidney and helps reduce water loss via the urine.
However, recent work suggests that specific adaptations for life in hot and dry desert climates may have evolved in desert birds. Tieleman and Williams (2000) compiled available values for basal metabolic rate (BMR), field metabolic rate (FMR) and field water flux (FWF), for 21 small bird species living in Old World deserts, and compared these data with the equivalent values for 61 species living in mesic habitats with higher rainfall and denser vegetation cover. The desert species included desert larks, sparrows and finches; the mesic species included owls, finches and sparrows, so there was a wide spread of groups. Two different methods of analysis were used, one for simple comparisons between the two groups, and the other based on phylogenetic contrasts involving the inclusion of phylogenetic relationships in the analysis. Significant differences in BMR and FMR between desert and mesic birds support evolution of reduced BMR and FMR in desert species. Low BMR and FMR might be expected to be an advantage for desert birds because of the associated lower energy demand, and lower release of metabolic heat, and hence lower TEWL, required for dissipating metabolic heat at high T a. Low FMR values can be linked to the habit in many desert species of resting in the shade or in burrows during the hottest part of the day (recall dune larks from Section 2.4). Values for FWF were significantly reduced in desert birds in comparison to mesic birds, but the difference was not related significantly to phylogeny.
Desert birds use panting for cooling, thereby incurring increased evaporative water loss. Female dune larks incubating their eggs pant during midday to regulate their own body temperature and hence that in their eggs. Desert grouse (Pterocles spp.), use gular flutter, a rapid vibration of the floor of the mouth that provides rapid evaporative heat loss with up to 2°C cooling in the mouth. Pterocles spp. can afford to lose water in this way, as these birds fly long distances every day to drink water from pools. Other birds such as desert larks do not show this behaviour and they may rely entirely on water obtained from their food, so they could not afford to lose so much water by evaporation.
Caution is advisable when using interspecific comparisons to support the view that physiological traits are adaptations. Physiological traits measured in species at different times of year or in different areas may vary, not because of genetic differences, but because of acclimatisation. A study of phenotypic flexibility of BMR and TEWL in 12 hoopoe larks (Alaemon alaudipes; Figure 35), captured from the Arabian desert provides a salutary example.

Two groups of six wild-captured larks were kept respectively at ambient temperatures of 15°C and 36°C, fed ad libitum and exposed to 12-hour day and 12-hour night regimes. After 3 weeks of acclimation, each bird was placed in a metabolic chamber at 35°C, a temperature within the thermoneutral range for the hoopoe lark. BMR was measured as the basal rate of oxygen consumption, and TEWL determined from water content of air expired from the chamber. The results are summarised in Table 5.
Initially there was no significant difference (P> 0.25) in the mean body mass of the two groups of larks. After 3 weeks of acclimation, mean body mass of the group acclimated at 15°C was significantly higher than that of the group acclimated at 36°C (P
| T a for acclimation | Body mass pre-acclimation/g | Body mass post-acclimation/g | BMR/kJ day−1 | BMR/kJ day−1 g−1* | TEWL at 35° C/g H2O day−1 | TEWL at 25° C/g H2O day−1 |
|---|---|---|---|---|---|---|
| 15°C | 41.3 ± 7.0 | 44.1 ± 6.5 | 46.8 ± 6.9 | 3.55 ± 0.60 | 3.11 ± 0.4 | |
| 36°C | 37.2 ± 4.7 | 36.6 ± 3.6 | 32.9 ± 6.3 | 2.23 ± 0.28 | 2.17 ± 0.7 | |
| P for difference between means | > 0.25 |
Activity 9
Compare the mean values for whole body BMR in the two acclimated groups of larks.
Answer
Larks acclimated at 15°C had a greater mean BMR, 46.8 kJ day−1, than the mean BMR, 32.9 kJ day−1, measured for birds acclimated at 36°C. The increase in BMR is statistically significant (P
The BMR of hoopoe larks acclimated to T a = 15°C approaches that reported for a temperate species, the woodlark (Lullula arborea): 49.4 kJ day−1.
Activity 10
You may argue that BMR expressed as kJ day−1 g−1 is likely to be the same for the two groups of hoopoe larks because body mass for the 36°C group is greater than that of the cold-acclimated group. Note down the values for BMR as kJ day−1 g−1 (see the empty column in the table) and state whether the BMR expressed per gram is still lower in the warm-acclimated larks.
Answer
Mean BMR value, 0.89, for the 36°C group is still lower when expressed as kJ day−1 g−1 than the equivalent mean value for the 15°C group, 1.06 kJ day−1 g−1.
Activity 11
Compare the mean values for TEWL in the two acclimated groups of larks.
Answer
Larks acclimated at 15°C had a greater mean TEWL at 35° C, 3.55 g day−1, than the mean TEWL 2.23 g day−1, measured for birds acclimated at 36°C. The increase in TEWL is statistically significant (PT a = 25°C.
Those larks acclimated to 15°C had significantly larger liver, kidney and intestine than larks in the 36°C group. Birds in the 15°C group consumed about 420 g food per day, more than three times as much as the 120 g food per day eaten by the birds kept at 36°C. The overall picture is that the hoopoe lark has high phenotypic flexibility, an advantageous feature for an animal living in a very variable environment. The environment of the Arabian desert has long periods of drought with scarce food resources available but unpredictable periods of rain temporarily increase food supply. The ability to minimise energy expenditure and requirement for water is important for survival of the birds. You may argue that the reduction in TEWL at 36°C derives from a lower BMR and therefore reduced respiratory evaporative water loss (REWL). Williams and Tieleman (2000) determined that REWL accounts for 31.7 per cent of TEWL at 35°C with cutaneous evaporative water loss (CEWL) accounting for the remaining 68.3 per cent. If it is assumed that the increase of 42.2 per cent in BMR in 15°C-acclimated birds results in an equal increase in REWL but no increase in CEWL, then TEWL would have increased by 13.4 per cent. The finding that TEWL increased by 59.2 per cent in the birds in the cold-exposure group indicates that they altered the permeability of their skin to diffusion of water vapour. Williams and Tieleman suggest that desert birds may reduce their CEWL by increasing their skin resistance, either by varying diffusion path length across the skin, or by altering the permeability of skin to water vapour. Diffusion path length can be reduced by vasodilation of subcutaneous capillary beds. Changes in lipid of the skin and increased epidermal thickness may reduce permeability of the skin to water in desert birds.
Studies on small desert birds suggest that lower BMR, FMR and TEWL are typical physiological responses to hot arid environments. Williams and Tieleman suggest that reduced BMR and TEWL may have evolved in desert birds, even though phenotypic adjustments in these physiological variables may be considerable, as demonstrated by their work on hoopoe larks. Whether reduced BMR and TEWL in desert birds results from physiological acclimation, phenotypic plasticity or an inherited feature is investigated further in Section 5. We should also be aware that phenotypic plasticity and the ability to acclimatise physiologically are under genetic control.