

3 Introducing the primates
One group of accomplished tree dwellers are the primates - a term you perhaps think of as synonymous with monkeys and apes. Monkeys and some apes display some of the most striking adaptations to tree-living.
250 or so species of primate exist today; most taxonomists group them into 13 families. All share a lengthy list of defining features, mostly related to the following broad categories:
Limbs and locomotion. The hands (and often the feet) are grasping, with mobile fingers and toes, generally with touch-sensitive pads at the tips. The first digit (the thumb or the hallux, i.e. big toe) is normally divergent (i.e. points outwards) and in many species can be swivelled to bring its tip into contact with other digits; in other words, it is opposable, to a greater or lesser degree. (Try this with your own hand to verify it.) Rather than a curved and rigid claw, at least some of the digits of primates have flat nails, making manipulation (e.g. of food) practicable. They have very flexible shoulder joints; hindlimbs are normally dominant in locomotion.
The senses. The eyes point forwards and are set close together. The fields of view of the eyes overlap which, together with a distinctive 'wiring' arrangement linking the eyes and the brain, imparts stereoscopic, '3-D' vision. In general, vision (like hearing) is more significant than smell, and colour vision is widespread. The face is flattened, with the muzzle (i.e. the area of skin around the nostrils) foreshortened; this anatomical change may, in part, be a reflection of the diminished importance of smell.
The brain. This organ is relatively large. The cerebral cortex - in primates, more often called the neocortex - is elaborately folded and complex in structure. This part of the brain is involved with the highly complex processes that include learning, reasoning and memory.
In all, up to 30 or so diagnostic features of the type described above have been identified for primates, though not all primates display each trait. The others include: their distinctive dentition, linked with a generally omnivorous diet; the structure of the ear; and reproductive features, such as small litter sizes and long gestation periods, relative to body size. Primates have an extended period of juvenile growth and their overall rates of growth and reproduction are generally low, which some biologists interpret as the price paid for a large brain. Brain tissue is metabolically a very expensive tissue to develop and maintain, so for large-brained mammals, less energy is available for growth and reproduction. The benefits of a large brain are very significant. For example, the elaborate social behaviour of primates is seen by many biologists to be as much a defining feature of primates as the types of anatomical feature just listed.
While monkeys and apes (and ourselves) may be the most familiar primates, they are far from the only members of this order. I'll focus on types of primates other than apes or monkeys, where again a tree-dwelling lifestyle is especially widespread. Such primates comprise three subgroups, all introduced in LoM:
the bush-babies (or galagos) of Africa (Section 4);
the lorises of Asia and the pottos of Africa (Section 4);
the lemurs of Madagascar (Section 5).
LoM refers to all such primates as prosimians [p. 230]; by contrast, monkeys and apes are termed 'anthropoids'. The term prosimian implies that these primates are in some sense 'forerunners of the monkeys' - historically monkeys and apes were referred to as 'simians'. More than likely, prosimians do more closely resemble the ancestral primates than do modern-day monkeys and apes, though thinking of them simply as primitive 'pre-monkeys' would be a mistake. As you'll see, they have complex and rich lifestyles and display sophisticated adaptations to tree dwelling.
LoM mentions one type of prosimian that stands apart from its relatives [pp. 243-245], the wholly carnivorous tarsiers, and attention is drawn to their rather anomalous taxonomic status. Indeed, most taxonomists now favour grouping tarsiers within the primate suborder that comprises the true monkeys and apes and urge the abandonment of the term prosimian. You're likely to encounter this preferred classification scheme in your wider reading and in the classification scheme featured in the Open University course from which this course originates (S182), so I'll outline it here.
The newly defined suborder Strepsirhini includes what DA refers to as prosimians, i.e. galagos, pottos, lorises and lemurs - but excludes the tarsiers. (The suborder takes its name from the features of the nose, in particular a naked area (the rhinarium) surrounding what are often referred to as 'comma-shaped' nostrils.) The muzzle of strepsirhines is commonly described as 'prominent, moist and glandular' and, as I've hinted, strepsirhines are thought, for a variety of reasons, to resemble more closely the ancestors of primates. Compared to other primates, they have smaller brains, a better sense of smell and longer snouts. As DA's comments imply, the tarsier's muzzle is different: 'fur grows almost to the edge of them [the nostrils] and surrounds them' [p. 245], i.e. the muzzle is hairy. This feature defines the suborder Haplorhini, together with the fact that the muzzle is less prominent. The haplorhines therefore include the tarsiers and the monkeys and apes.
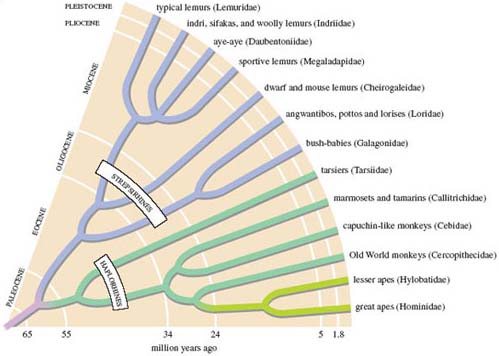
This revised classification is meant more accurately to reflect the presumed evolutionary history of primates. Figure 2 provides such an evolutionary tree, plotting the different lineages that evolved from the presumed common ancestor of primates, stretching back into the Paleocene. You can see a divergence from this ancestor into the two branches reflecting the suborders just introduced, the differences in colour evident as they split. Continued branching of each of these evolutionary lines, at different periods in the geological past produced - perhaps as much as 100 million years or so on from the ancestral primate - the range of 13 families that comprise today's primates. But for consistency with LoM, I'll refer to the two suborders of primates as prosimians and anthropoids, rather than using the haplorhines and strepsirhines division of Figure 2.
I've already drawn attention to the distinctive hands and feet of primates. shows some of this anatomical diversity in primates, including representatives of the monkeys and apes. The hand of a tree-shrew, i.e. a non-primate, is shown for contrast (Figure 3a); this climbing mammal is not particularly well adapted for living in trees (some species of tree-shrew are almost entirely ground-dwelling). Tree-shrews have claws on all their fingers and their thumb - and the thumb is hardly different from the other digits. The digits are able to do little more than spread out or converge, rather than span and grip. In contrast, the feet and hands of the primates shown in Figures 3b-j generally have nails, which are straighter and less robust than claws. They also have tactile pads and a more manoeuvrable thumb and toe - features that are used to good effect in the tree-climbing species illustrated here. Figure 3 reveals the great anatomical diversity of primate hands and feet and a widespread ability to grasp, i.e. they are very often prehensile, which is reason enough for us to come back to the figure later in this course.
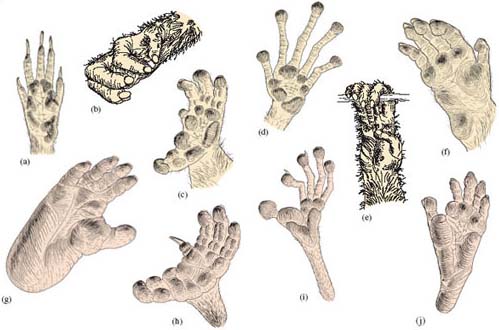
In Figure 3 (a) is the paw of a tree-shrew, for contrast. (b) shows the precision grip of the chimpanzee, reflecting their higher degree of manual dexterity. (c) and (h) are the hand and foot, respectively, of a potto; (d) and (i) are the hand and foot, respectively, of a tarsier. (e) is the hand of a gibbon; the thumb is short, highly opposable and distant from the remaining digits, which are slender and well suited to grasping. The arm-swinging form of locomotion of gibbons, which is called brachiation. The relatively long forelimbs, together with anatomical adaptations that increase the mobility of the shoulder, forearm and wrist, contribute to this very efficient form of locomotion, ideally suited to an environment where handholds are irregular and widespread. (f) is the hand of a macaque monkey, which is adapted for walking with the palm flat on the ground. (g) and (j) show the foot of a gorilla and baboon, respectively, both largely ground-dwelling primates.