

1.4.4 Hibernation in mammals
Many animals become inactive for periods of varying duration during the winter and there is a diversity of terms used to describe this state, including: sleep, torpor, dormancy, lethargy and hibernation. The word hibernation is often used loosely to refer to general inactivity but, in biology, it refers to a specific phenomenon, sometimes called 'true hibernation'. Hibernation is defined as the condition of passing the winter in a resting state of deep sleep, during which metabolic rate and body temperature drop considerably. It occurs only in certain mammals and one bird species, the poorwill (Phalaenoptilus nuttallii), a North American relative of the nightjar.
The phenomenon of hibernation is one reason why the term homeothermy is going out of fashion, to be replaced by endothermy, because maintaining a stable body temperature is the very opposite of what hibernators do. Instead, body temperature falls, from around 38°C, to about 1°C above ambient temperature, which is often close to 0°C. At the same time, a hibernator's metabolic rate falls to as little as 1% of its normal value. The heartbeat becomes slow and irregular and breathing rate also slows.
Hibernation is an active process, that is, it is a state which animals enter into, not in response to immediate external conditions, but to internal stimuli. Some species are remarkably precise and predictable. For example, the arctic ground squirrel (Spermophilus undulatus) enters hibernation between 5 and 12 October and emerges between 20 and 22 April, regardless of the weather on those dates. This behaviour is in contrast to other winter states such as torpor or lethargy which are immediate responses to current conditions. Brown and black bears, for example, are lethargic during very cold periods but are otherwise active in the winter. A feature of hibernation that distinguishes it from other kinds of winter inactivity is that hibernators can arouse themselves spontaneously and are not dependent on external conditions, such as warm temperatures, to do so. The arctic ground squirrel is described as an obligate hibernator because it hibernates every winter. There are some mammals that are categorised as facultative hibernators, entering hibernation in response to very cold weather and poor food supply. The North American pocket mouse (Perognathus californicus) is a facultative hibernator.
True hibernation only occurs in relatively small mammals, though not all small mammals living in temperate habitats hibernate in winter, as we have seen. The largest mammal to hibernate is the marmot, which weighs about 5 kg. There are several reasons why larger mammals do not hibernate. Firstly, they would warm up too slowly and therefore use too much energy. Secondly, they have a smaller surface area to volume ratio and so can conserve body heat better than smaller species. Finally, they are better able to carry a thick coat (Section 1.3.2) and sufficient adipose tissue to last through the winter. Hibernators are mainly found in the orders Rodentia, Chiroptera (bats) and Insectivora. The hedgehog (Erinaceus europaeus) is an example of a hibernating insectivore; in Britain, it hibernates from October/November to March/April. Note that although hedgehogs are in the order Insectivora they do not just eat insects!
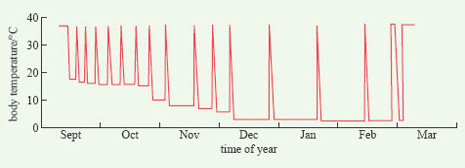
The physiological features that are characteristic of hibernation are not maintained throughout the winter. Rather, the animal wakes up at intervals, its temperature and metabolic rate increasing to near-normal levels (Figure 15). The function of this periodic arousal is not wholly clear. Some species, such as the chipmunks (genus Tamias) eat from stored food reserves during arousal periods, but many others do not. Most species urinate and defecate, move about and change their position, suggesting that arousal provides an opportunity for various essential physiological processes to be performed and to prevent the animal becoming moribund. From detailed measurements of Richardson's ground squirrel (Spermophilus richardsonii) in the laboratory, it has been calculated that, during the relatively brief periods of arousal (Figure 15), an individual expends 83% of all the energy that it uses up during the entire hibernation period.
Hibernation requires internal energy reserves in the form of adipose tissue and hibernators typically feed intensively prior to winter, building up their fat stores. Some species, such as the edible dormouse (Glis glis), switch to a carbohydrate- and lipid-rich diet, e.g. seeds, at this time. A characteristic of hibernating mammals is that they possess larger quantities of a particular kind of adipose tissue called brown adipose tissue (BAT). This tissue gets its name from its dark colour, which is due to the larger numbers of blood capillaries that permeates it and the high concentration of mitochondria within the cells. BAT is rich in mitochondria with special properties that enable it to oxidise fatty acids and/or glucose to produce heat very rapidly. BAT deposits are found around some internal organs and between the shoulder-blades of hibernators and their function is to generate body heat very rapidly, especially during periods of arousal.
Hibernation might seem to be a safe, and rather agreeable way to spend the winter but, for some species, it is fraught with danger. For Belding's ground squirrels (Spermophilus beldingi) living at high altitude in Tioga Pass, California, hibernation lasts 7-8 months. Two-thirds of all juveniles, hibernating for the first time, and one-third of adult animals die during hibernation. Some die because their fat reserves run out before the end of hibernation; others are dug up and eaten by predators.
Some mammals spend the winter in groups, huddled together during periods of dormancy, and so conserve body heat. North American raccoons (Procyon lotor), for example, spend dormant periods in communal dens. Many species of bats hibernate communally. During hibernation, the body temperature of some bats can fall below 0°C. In the autumn, they build up fat reserves that represent as much as a third of their total mass. During the winter, bats arouse themselves from hibernation to excrete and sometimes also to move to a new roost. A critical factor for hibernating bats is that roosting sites have high humidity and some populations have to migrate quite large distances to find suitable places, such as caves and hollow trees.
For some mammals, hibernation is closely associated with other important activities, notably reproduction and dispersal. Consequently, energy reserves may have to support more than one activity. For example, brown bears living at northern latitudes mate in the autumn and give birth to their cubs during winter lethargy. Edible dormice and some bats mate immediately after the end of hibernation. (In some species of bats, males wake up first and mate with the females before they have woken up!) The link between what animals do in winter and their reproductive cycles was discussed in Section 1.2.3.
Natal dispersal is the permanent departure of an individual from its place of birth, usually at the end of the breeding season. It is an important part of the life history of many animals, especially mammals, and tends to be sexually dimorphic, males dispersing further than females. Natal dispersal is potentially both hazardous and energetically expensive; dispersing animals tend to be vulnerable to predators and, being on the move, have little time to feed. Dispersal therefore requires internal energy reserves in the form of fat, the very same reserves that they later need to survive the winter. There may thus be a trade-off in the allocation of energy reserves to dispersal and to hibernation.
Scott Nunes of Michigan State University has studied dispersal and hibernation in Belding's ground squirrels in a locality where hibernation lasts for eight to nine months of the year. Young males typically disperse after the summer breeding season but show much variation in the extent to which they do so, with fatter males being more likely to move out of the natal area. In years when breeding is delayed, dispersal is inhibited; instead, young males remain near the natal area, building up their fat reserves prior to hibernation. The findings of this study are summarised in Figure 16.
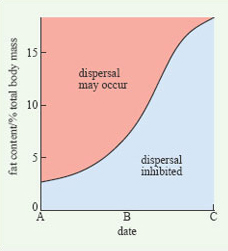
If young males are ready to disperse early (date A in Figure 16), they leave with relatively small fat reserves. In this situation, they have time to travel further to a new area and then build up fat reserves before hibernation. If dispersal is delayed (dates B and C), males do not disperse unless they have built up a threshold level of fat reserves; the value of this threshold increases as winter approaches. In other words, the trade-off between dispersal and hibernation is resolved by hibernation suppressing dispersal, unless an animal exceeds a certain fat level. After date C, dispersal is inhibited regardless of the size of the fat reserves.