

1 The desert climate: An introduction
If you have visited a desert you will have noticed the sparse plant cover, or in certain sandy deserts, the almost complete absence of plant life. The low productivity of deserts derives from their defining feature, which is aridity. Scarcity of water restricts the diversity and amount of plant cover, and in turn the diversity and abundance of animals. However, if you were visiting one of the American deserts after rains, you would be rewarded by the sight of the desert ‘in bloom’, as vast swathes of annual plants such as the Mojave aster (Xylorhiza tortifolia) and sand verbena (Abronia villosa) flower simultaneously. You might catch sight of insects such as beetles and locusts, and vertebrates including lizards and occasionally mammalian herbivores, such as gazelle, in African deserts and oryx in the Arabian desert.
Hot deserts located 15°–25° north and south of the Equator have daytime sunshine all year round (Figure 1). The persistent descending air and stable high pressures create the hot climate. While daytime temperatures can be as high as 45°C, night-time temperatures may be close to freezing as heat is lost by radiation into the clear night skies.
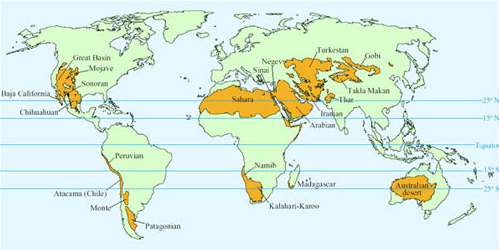
Aridity of deserts has three main causes. The deserts in parts of North and South America are arid because they are located on the leeward side of mountain ranges in rain shadow. Rain falls as moisture-laden air rises up the mountains, so that the air is dry by the time it reaches the leeward side. The Gobi and Turkestan deserts lie in the centre of a large continent and their lack of rainfall is because they are a long distance from the sea. The Sahara (Figure 2) and Arabian deserts are arid because of persistent large high-pressure masses of dry air that prevent penetration of rain-bearing storm systems. A popular concept of deserts is based on the extreme Sahara, where huge areas of sand dunes support little, if any, plant growth. Certain animals such as camels (Camelus spp.; Figure 3), Dorcas gazelle (Gazella dorcas; Figure 4) and oryx (Figure 5) that survive there by browsing on the sparse plant life and drinking very little or no free water, are regarded as typical desert species.




In fact the picture is much more complex; the environment of each desert is unique, and depends on the interaction between T a, precipitation, relative humidity and wind. A useful classification is that of Meigs (1953), who defined deserts according to aridity (Table 1). The aridity of a desert is determined not just by precipitation but also by the evaporation and transpiration of plants. In order to simplify classification of arid and semi-arid areas, various types of aridity index have been devised. De Martonne's aridity index has been used widely and is calculated from the formula:

I a = aridity index
P = mean annual precipitation/mm
T = mean annual temperature/°C
Note that values for average precipitation can be misleading because in arid deserts, especially hyper-arid desert, there are many years that have no rainfall at all. Certain coastal deserts such as the coastal strip of the Arabian Peninsula obtain part of their annual precipitation from thick fog, caused by cold sea breezes that increase humidity sharply. Subdivisions of aridity index are roughly the same as those for average rainfall.
| Rainfall/mm yr−1 | Aridity index | Aridity | Examples |
|---|---|---|---|
| < 25 | < 5 | Hyper-arid | Namib; Arabian |
| 25–200 | 5–20 | Arid | Mojave |
| 200–500 | 20–50 | Semi-arid | Parts of Sonoran |
All deserts have a wide range of T a, but mean annual temperatures vary from desert to desert. Deserts can be defined as hot, mild, cool or cold (Table 2), but the reality is a continuum of desert climates rather than a set of desert climates each with a well-defined rainfall and T a range.
| Climate | % of deserts | Examples | Mean T a coldest month/°C | Mean T a warmest month/°C |
|---|---|---|---|---|
| Hot | 43 | Central Sahara; central Australian | 10–30 | > 30 |
| Mild | 18 | Kalahari-Karoo, Chihuahuan | 10–20 | 10–30 |
| Cool | 15 | Mojave, Namib | 0–10 | 10–30 |
| Cold | 24 | Gobi | < 0 | 10–30 |
Deserts may have seasonal climates, with winters being much colder than the summers. The Sonoran desert covers about 260 000 km2 and spans the western part of the Mexican state of Sonora, southwest Arizona, southeast California and Baja California. Average rainfall is 120–300 mm yr−1, with the rain falling in two seasons. Storms from the North Pacific bring gentle rain from December to March, and surges of wet tropical air bring in rain storms from May to September. Winter temperatures are cool, averaging 13°C, and summers are extremely hot, reaching 40°C on average, but peaking at 50°C. Ambient temperatures can vary by as much as 40°C in a day. It is also important to appreciate that there are variations in climate, topography and vegetation within a desert. The Lower Colorado River Valley region of the Sonoran desert (Figure 6) is the driest hottest area, where annual rainfall may be < 50 mm and summer daytime temperature 50°C, with surface temperatures reaching 82°C; according to Meigs’ classification this desert would be classed as arid hot. Vegetation there consists of drought-tolerant shrubs and succulent cacti; following winter rainfall there is rapid growth of annual plants, which cover the ground in a mass of blooms. Within the Lower Colorado River Valley there is a large area of sand dunes forming a huge sand sea. In contrast, the Arizona Upland region has an annual rainfall of 70–300 mm, making it an arid or semi-arid desert according to Meigs. There is abundant vegetation including trees such as ironwood, blue paloverde and cat claw acacia. Cacti include 12 species of cholla (Opuntia spp.) and also saguara cactus (Figure 7). Scrub plants include desert saltbush and creosote bush. Winter annuals, e.g. California poppies, bloom in profusion after rain.


The Mojave desert spans the transition between the Sonoran and Great Basin deserts and extends throughout southeastern California, and parts of Nevada, Arizona and Utah, occupying about 100 000 km2. Summers are hot and windy, but during the winter, temperatures can dip to below freezing. The Mojave desert is arid, with only about 130 mm rainfall per year in a winter rainy season, but the rains fail in some years. The plant life in the Mojave desert comprises Yucca species, including the joshua tree, big sage brush, bladder sage and creosote bush and at least 200 other endemic species.
Despite the variations in the environment of different deserts, it is correct to say that all desert animals have to cope with water shortage, and animals living in hot deserts cope with extremely high daytime T a. Physiological problems linked to high T a are those associated with hyperthermia. Mammals and birds have an optimal core T b of around 38°C, and many species cannot tolerate increases > 2°C or so. The denaturation of crucial proteins, such as enzymes, begins at around 40–42°C, so hyperthermia also creates physiological problems for ectotherms. Daytime temperatures in desert environments can be much higher than the optimal T b, e.g. up to 56°C in Death Valley, California. Homeotherms subjected to heat stress use a suite of physiological mechanisms for cooling the body, which we will explore in later sections. Evaporative cooling is the most effective way for an animal to lower T b, yet if water is in short supply, dehydration is a serious problem, and the use of evaporative cooling is restricted. Behavioural mechanisms play an important role in cooling the body, both in desert ectotherms such as lizards, snakes and amphibians, and endotherms, such as birds and mammals.
Summary of Section 1
Despite deserts being diverse, they all have aridity in common as their salient climatic feature. Classification systems attempt to group deserts in terms of their aridity or mean annual temperatures. Major physiological problems for animals living in deserts include hyperthermia due to intense daytime solar radiation, and also hypothermia at night when desert T b can be below freezing. The shortage or lack of drinking water in deserts means that evaporative cooling cannot be used freely for physiological thermoregulation.