

2.4 Behavioural strategies of evaporators
Willmer (2000) defines evaporators as animals that depend on sufficient water intake to enable them to cool T b by evaporation. Few of these species can survive in deserts, and those that do either live on the edges of deserts where they can access water, or have behavioural and physiological adaptations that reduce reliance on evaporative cooling. So for evaporators, evasion may be an important part of their thermoregulatory strategy. Evaporators include medium-sized mammals such as jack rabbits, dogs, foxes, and also desert birds such as larks.
The jack rabbit (Lepus californicus; Figure 21) is a hare, living in the Sonoran and Mojave deserts. Jack rabbits do not burrow, although they are quite small, weighing about 2 kg.

A jack rabbit would need to lose at least four per cent of its body mass per hour to thermoregulate by evaporation. There is little or no free water around; water is obtained from the diet, green plants, including cacti in the summer. Knut Schmidt-Nielsen's work (1967) showed that behaviour is important for the jack rabbit's survival. During the hottest part of the day the animal chooses a shaded depression in the ground, often in the lee of a bush, in which it crouches (Figure 22).
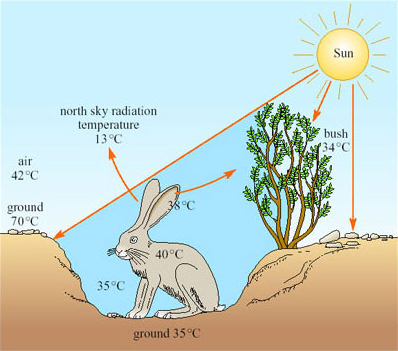
The bottom of such a depression has a much lower temperature than that of the rest of the surface, the hot desert wind and much of the radiation passing over the animal's head. From its sheltered position, the jack rabbit's large radiator-like ears can be exposed, not directly to the Sun, but to a clear blue sky. The radiation temperature of the north sky at midday is only 13°C so if the ears, which are richly vascularised, have a temperature of 38°C, and have a surface area of 400 cm2, are directed towards the sky, they can radiate about 13 kJ h−1, which is about half of the animal's metabolic heat production. The jack rabbit forages during the night.
The kit fox (Vulpes macrotis; Figure 23) lives in the Sonoran, Mojave and Great Basin deserts in southwestern USA. Kit foxes have very large ears, which are thought to provide an increased surface area for cooling the body.

They are carnivores, and hunt at night, preying on kangaroo rats, tortoises and jack rabbits, and occasionally catching ground-nesting birds, reptiles and insects. They reduce evaporative water loss by spending the day in underground dens, emerging at sunset to begin hunting. The physiological importance of dens for desert foxes should not be underestimated. By remaining in the den during the day, a desert fox reduces drastically the need for panting, a mechanism used by foxes and dogs for cooling the body by evaporative water loss (Section 3.3).
A few species of small birds live in the most extreme deserts. Dune larks (Mirafra erythroclamys; Figure 24) are the only birds that live year round in the Namib sand sea, one of the driest regions of the world. Dune larks feed on insects and spiders, which they collect during the day, while walking over the sand surface; they also peck insects from just below the sand surface. In winter the birds feed on seeds blown in from adjacent grass land. The scarcity of water in the Namib sand sea means that dune larks drink rarely and the birds rely on water in their food and on metabolic water. Birds do not sweat, but they use both cutaneous and respiratory evaporative water loss for cooling the body. During the hottest part of the day, from around 12.00 to 15.00 h, dune larks seek shade and stand still. Presumably this behaviour helps the birds to cool T b and reduces evaporative water loss.

Williams (2001) used taxidermic mounts to determine operative environmental temperature, T e, for the birds. T e is the temperature that an animal would reach in the environment if it was biologically inactive, i.e. only the physical characteristics of the animal are taken into account. It is defined, in physical terms, as the temperature of a black body of uniform temperature, in an identical situation to that which the animal occupies, with the same values for conduction, convection and radiation. As the definition is purely physical, it is possible to make models of animals and to use them to measure T e experimentally. Figure 25 shows three examples of daily profiles for a model of a dune lark made from a copper cast of a bird covered in plumage.
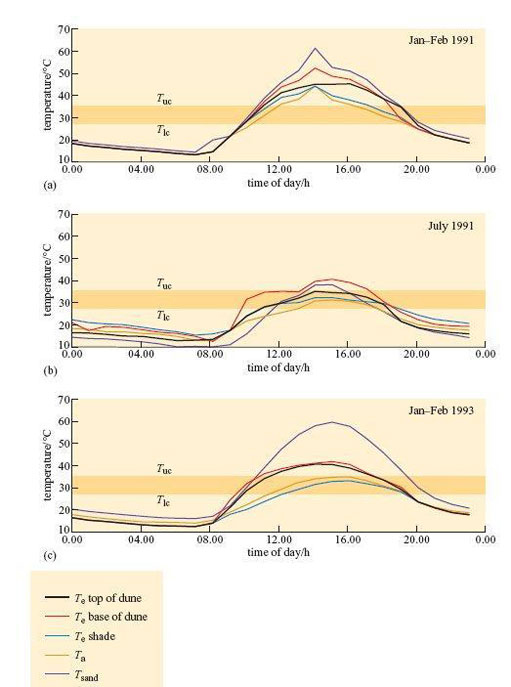
Figure 25b shows that during July 1991 (winter in Namibia) mean T e shade did not exceed T uc (35.1°C) for the larks. The results suggest that in winter, the strategy of finding a shady spot during the hottest part of the day lowers T b sufficiently, so there is no need for physiological cooling, in particular evaporative water loss, for maintaining T b.
Activity 5
Do the results shown in Figures 25a and c suggest that dune larks do not need to use evaporative cooling to maintain T b in the summer? What is the main advantage of resting in the shade for the dune lark? Identify one disadvantage.
Answer
Even in the summer, T e in shade is significantly lower than T e in sun. In Jan–Feb 1991, at midday, all mounts exposed to the sun reached a T e of 46–50°C, higher than T uc of 35.1°C. T e in the shade was significantly lower. In summer 1991, mounts exposed to full sun reached T e values of 40–50°C from about 12.00–20.00 h, whereas those in the shade peaked at 40–45°;C from about 12.00–16.00 h. In summer 1993, T e for mounts in the shade never exceeded T uc. For the dune lark, the simple strategy of standing in shade during the heat of the day provides significant cooling, even in a particularly hot summer like that of 1991. It is likely that by standing in shade, the need for evaporative cooling must be reduced at least.
However, the disadvantage of this strategy is that while standing in the shade, dune larks cannot forage, so the bird has to find a balance between the need for food and the necessity of avoiding excessively high T b.
While desert animals classed as ‘evaporators’ could use evaporative cooling for maintaining T b at high T a, the need for this is avoided by simple behavioural strategies. Nocturnal foraging and daytime use of dens, burrows and shade for cooling reduce the need for physiological cooling by evaporative water loss, thereby conserving water.