

6 Phylogeny and cladistic analysis
In Section 3.3 the point was made that many physiologists consider that desert birds are successful because of their avian physiology, not because of any specific adaptations. While Williams and Tieleman's research on hoopoe larks demonstrated that desert species are capable of flexibility in metabolic rate and evaporative water loss, it suggested that adaptation is important too. The selective advantages of lowered BMR and TEWL for desert birds include reduced energy demand, and lower production of metabolic heat reducing the need for EWL. Both BMR and TEWL appear to be at least partly genetically determined, so it is likely that they are subject to natural selection. We have seen from the hoopoe lark study in Section 3.3 that phenotypic flexibility may account for differences between populations of the same species. Phenotypic plasticity (Section 3.3) may also provide confounding variables for such studies.
The power of the comparative approach in which a physiological feature in a range of related species is studied can be appreciated from the study of Peromyscus species outlined in the previous section. Studies of clades comprising related genera within a family can be more powerful still. Irene Tieleman et al. (2003) tested the hypotheses that both BMR and TEWL are reduced along an aridity gradient within a single family of birds, Alaudidae (larks). Their study also investigated the role of phylogenetic constraint, the situation where similar traits seen in closely related species may be there not because they are adaptive, but because they were inherited from a common ancestor. The researchers chose larks for the study, because larks live in a vast geographic area, covering three continents, with species found in a wide range of environments, from hyper-arid desert to mesic grassland. Nevertheless, all larks eat a similar food, seeds and insects, so diet was not a confounding variable in this study.
Aridity is directly related to primary productivity, and is therefore a sound measure of the selection pressures that animals experience with increasing aridity. These selection pressures include decreased food and water availability and increasing air temperatures. The researchers investigated eight species, and also used data in the literature for six other species for the study. BMR and TEWL were measured at night using standard respirometry and hygrometry methods. The aridity index selected, log Q, was appropriate to the habitat of the birds in this study. Log Q is low in hot dry deserts and high in relatively cool wet areas.
Table 9 summarises the results for the 14 lark species. Analysis showed that both body mass and aridity had significant effects on BMR (P< 0.0001).
| Species | Body mass/g | BMR/kJ day−1 | TEWL/g H2O day−1 | Annual precipitation/mm | Log Q |
|---|---|---|---|---|---|
| Grey-backed finchlark (Eremopterix verticalis) | 15.1 | — | 1.31 | 33.0 | 1.76 |
| Stark's lark (Eremalauda stark) | 15.6 | — | 1.31 | 57.2 | 1.76 |
| Desert lark (Ammomanes deserti) | 21.5 | 20.1 | 1.60 | 89.6 | 1.78 |
| Dunn's lark (Eremalauda dunni) | 20.9 | 24.7 | 1.69 | 89.6 | 1.78 |
| Hoopoe lark (Alaemon alaudipes) | 36.9 | 32.8 | 2.59 | 89.6 | 1.78 |
| Black-crowned finchlark (Eremopterix nigriceps) | 15.2 | 16.5 | 1.34 | 209.1 | 2.24 |
| Crested lark (Galerida cristata) | 31.2 | 32.2 | 2.44 | 209.1 | 2.24 |
| Calandra lark (Melanocorypha calandra) | 50.6 | 49.5 | 3.03 | 250.0 | 2.26 |
| Horned lark (Eremophila alpestris) | 26.0 | 28.6 | 2.08 | 309.9 | 2.41 |
| Lesser short-toed lark (Calandrella rufescens) | 23.6 | 31.6 | — | 281.0 | 2.59 |
| Short-toed lark (Calandrella brachydactyla) | 24.0 | 35.6 | — | 281.0 | 2.59 |
| Spike-heeled lark (Chersomanes albofasciata) | 25.7 | 29.1 | 3.33 | 420.4 | 2.60 |
| Skylark (Alauda arvensis) | 32.0 | 62.4 | 3.17 | 750.0 | 3.20 |
| Woodlark (Lullula arborea) | 25.6 | 49.4 | 2.41 | 750.0 | 3.20 |
Tieleman et al.'s analysis of the data in Table 9 supports their view that BMR of larks increases as the environment becomes more mesic (Figure 46a). Also, TEWL of larks decreases as the environment becomes more arid (Figure 46b).
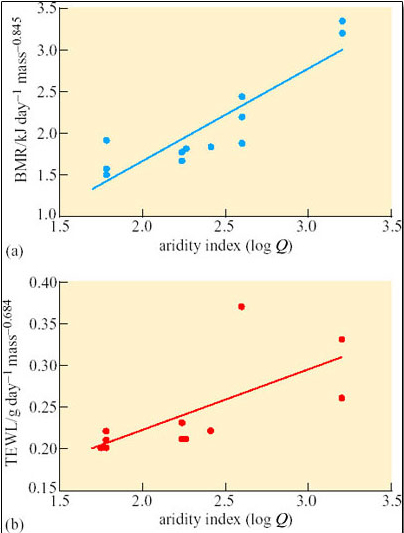
Tieleman et al.'s study involved the comparison of two traits across lark taxa, BMR and TEWL. The statistical analyses of the data in Figures 46a and b assumed phylogenetic independence, but the researchers were aware that the positive correlation obtained between BMR and TEWL with aridity could be explained by phylogenetic autocorrelation, a relationship derived from the phylogeny itself, rather than adaptation.
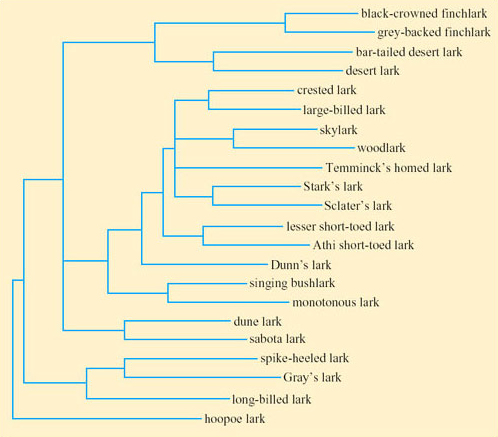
The researchers constructed a cladogram for 22 species of lark. DNA samples were extracted from blood or tissue samples, and polymerase chain amplification was used to amplify the cytochrome b gene (975 base pairs) and the 16S rRNA gene (566 base pairs). DNA was sequenced for each species and statistical analysis carried out to determine the most probable evolutionary tree for the clade. The cladogram (Figure 47) indicates a number of clades within the lark family. The lengths of the branches in the cladogram indicate phylogenetic distance between the clades.
Activity 19
Compare the lengths of the terminal branches of the cladogram with those of the internal (initial) branches.
Answer
The terminal branches are long in comparison to the short internal branches.
Activity 20
Suggest an outgroup within the cladogram and justify your choice.
Answer
The position of the hoopoe lark suggests that it may be an outgroup. The hoopoe lark is placed at the bottom of the cladogram suggesting that this species might be representative of the common ancestor.
Tieleman et al. interpret the short internal branches, coupled with long terminal branches, as suggesting that lark species underwent rapid adaptive radiation and have lived for a major part of their evolutionary history in diverse habitats. It is possible that variation in TEWL and BMR relates to phylogenetic distances, so that the more closely related genera within a clade have similar TEWL and BMR. For example, if variation in BMR could be explained simply by phylogenetic distance, members of one clade within the lark family would have similar values for BMR, in contrast to members of two other clades, which might have lower and higher values, respectively. In that case, BMR would be correlated with phylogeny itself, i.e. phylogenetic autocorrelation, which may be attributable to phylogenetic constraint or to ecological factors.
There was a significant phylogenetic effect in the data linking mass-corrected BMR with aridity.
Specialised statistical analysis ruled out phylogenetic autocorrelation as the sole explanation of the relationship between BMR and aridity. Since the BMR of larks decreases along a gradient of increasing aridity, Tieleman et al. therefore argued that this relationship is due to adaptation rather than phylogenetic constraint.
Mass-corrected TEWL correlated with aridity with no significant influence of phylogeny.
The decreases in BMR and TEWL with increasing aridity in larks suggest that BMR and TEWL have adaptive significance and that natural selection is the relevant process.
Summary of Section 6
A cladistic analysis of 14 lark species (family Alaudidae) investigated the hypotheses that BMR and TEWL are reduced along an aridity gradient. Although larks are found over a huge geographic area, all species have similar diets so diet was not a confounding variable. The researchers collected data on body mass and BMR and calculated the aridity index for the environment of each species. Analysis of the data showed that aridity had a significant effect on both BMR and TEWL. The shape of an evolutionary tree for the clade suggests radiation of larks soon after their origin. No correlation was found between mass-corrected TEWL and phylogeny, suggesting that correlation between TEWL and aridity is not due to phylogenetic constraint. Statistical analysis eliminated phylogenetic constraint as an explanation for the relationship between BMR and aridity although mass-corrected BMR correlated with both aridity and phylogeny.