
1 General principles of signal transduction
1.1 Introduction
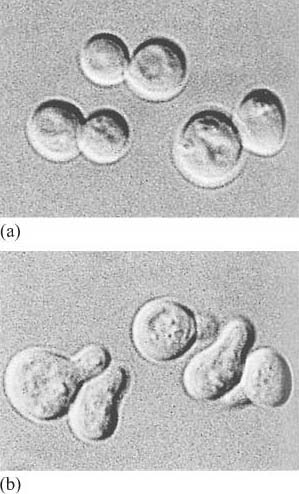
The fundamental principles of signalling can be illustrated by a simple example in the yeast S. cerevisiae (Figure 1). In order to sexually reproduce, a yeast cell needs to be able to make physical contact with another yeast cell. First, it has to ‘call’ to yeast cells of the opposite mating type. It does this by secreting a ‘mating factor’ peptide, an extracellular signal, which can also be called an ‘intercellular signal’. Yeast mating factor binds to specific cell surface receptors on cells of the opposite mating type, and the signal is relayed into the target cell via a chain of interacting intracellular signalling molecules, which switch from an inactive (Figure 2a) to an active state (Figure 2b). Signalling molecules are said to be upstream or downstream of other components of the pathway (this terminology should not be confused with that used to describe the structure of genes in relation to transcription). Ultimately, signalling molecules activate target effector proteins (an effector in this context is a molecule that carries out the cellular response(s) of the signalling pathway). In the yeast, signal transduction to mating factor ultimately stops the target cell proliferating, and induces morphological changes which result in the formation of protrusions towards the cell that releases the mating factor (Figure 1b). The morphological changes are a response to the signal. The two cells can then make physical contact with each other, and mating can ensue.
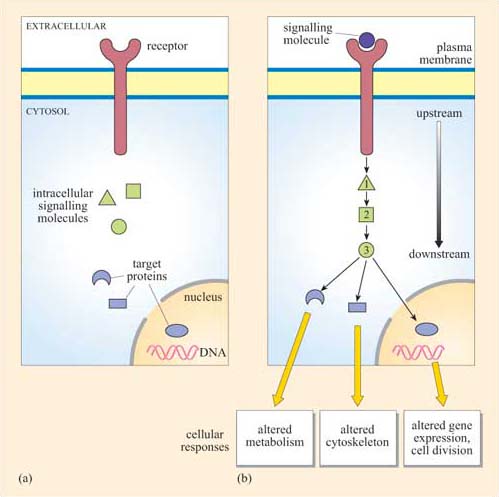
Which signal pathway molecule(s) can be said to be upstream and downstream of molecule 2 in Figure 2b?
The receptor and intracellular signalling molecule 1 are upstream; both intracellular signalling molecule 3 and the target proteins are downstream.
Signalling in multicellular organisms is a complex process, in which many millions of highly specialized cells may need to act in a coordinated fashion. Cells may need to respond to several signals at once, and different cells may need to respond to the same signal in different ways. All this is made possible because the mechanism for detection of a signal is not directly coupled to the response, but is separated by a chain of signalling events, such as that shown in outline in Figure 2b. This principle allows signalling systems to be highly flexible. Examples of this flexibility are:
the same type of receptor can be coupled to different signalling pathways in different cell types;
the signal can be amplified (or damped down) as it travels along the signalling pathway;
it can switch on multiple pathways, leading to several cellular responses in diverse regions of the cell;
information can be processed from several different receptors at once to produce an integrated response.
Most of this is made possible by protein–protein interactions and protein regulatory mechanisms.
Despite this complexity, the basic model of signal transduction set out in Figure 2 holds true for most intracellular signalling pathways across species, and often the signalling molecules themselves are highly conserved. For example, there is a high degree of homology between the major proteins in the yeast mating factor signalling pathway and the human mitogen-activated protein (MAP) kinase growth signalling pathway.
A mitogen is an extracellular molecule that induces mitosis in cells.
In this course, we shall guide you through the signalling network, firstly by introducing you to the kinds of molecules involved in signal transduction and the general principles employed by cells. Then we shall go into greater detail to show you exactly how the key molecular players operate including receptors and intracellular signalling molecules. In the final section, we shall consider specific examples of signal transduction pathways regulating cellular responses involved in glucose metabolism in different cell types.