
1.5 Signal transduction mechanisms
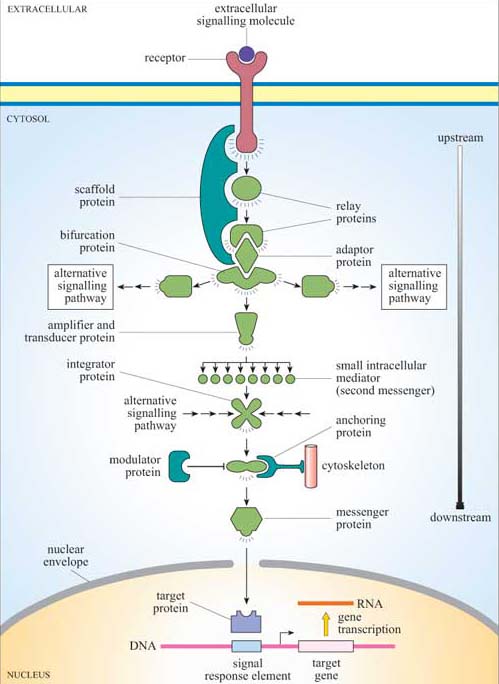
Signalling information has to be transmitted from the receptor in the plasma membrane across the cytoplasm to the nucleus (if gene transcription is the response), the cytoskeleton (if cell movement, or another change to cell morphology, is the response), or various other subcellular compartments. The transmission of a signal must occur in a time-frame appropriate for the cellular response. So, signal transduction needs to take place over both space and time. We have already described a simple signalling model (Figure 2), where a chain of intracellular mediators successively activates the next in the chain until the target is reached. In reality, of course, it is rarely a simple chain, but a branching network, allowing for integration, diversification and modulation of responses (Figure 6). The branched molecular network of activation (and deactivation) of signalling molecules linking receptor activation to the intracellular targets is referred to as a signal transduction pathway (or cascade).
Intracellular signalling molecules have particular properties that allow control of the speed, duration and target of the signal, and may be categorized according to these properties. Broadly speaking, intracellular signalling molecules can be divided into two groups on the basis of molecular characteristics, second messengers and signalling proteins.
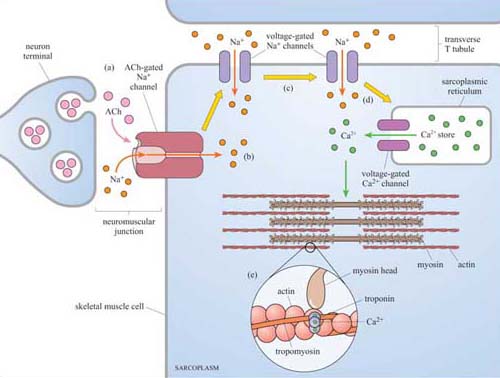
Second messengers are small readily diffusible intracellular mediators, whose concentration inside the cell changes rapidly on receptor activation; in this manner, they regulate the activity of other target signalling molecules. The calcium ion, Ca2+, is a classic example of a second messenger, being released in large quantities in response to a signal (so amplifying the signal) and diffusing rapidly through the cytosol. Ca2+ ions can therefore broadcast the signal quickly to several distant parts of the cell. For example, Ca2+ ions mediate and coordinate contraction of skeletal muscle cells (Figure 7). In general, if a rapid, generalized response is necessary, a second messenger is likely be prominent in the signalling pathway.
Other water-soluble second messengers such as cAMP and cGMP act similarly to Ca2+, by diffusing through the cytosol, whereas second messengers such as diacylglycerol (DAG) are lipid-soluble, and diffuse along the inside of the plasma membrane, in which are anchored various other key signalling proteins.
Second messengers were the first intracellular signalling molecules to be identified; they were so named because hormones or other extracellular signalling molecules were considered the ‘first messengers’. However, the term ‘second messenger’ seems somewhat outdated, since a signalling pathway can easily involve a sequence of eight or more different messengers, and the ‘second messenger’ in question could well actually be acting as, say, the fifth messenger.
Signalling proteins are the large intracellular signalling molecules that generally, but not exclusively, function by activating the next signalling protein in the signal transduction cascade, or by modifying the concentration of second messengers.
Proteins are much larger and generally less mobile than small water-soluble second messengers, so they are not so useful for the rapid dissemination and amplification of a signal. However, proteins are capable of interacting in a highly specific manner with other proteins, they exhibit binding specificity for ligands and for recognition motifs on other molecules, and their activity can be regulated, for example by allosteric regulation and by phosphorylation. They are therefore able to perform rather more sophisticated signalling roles than water-soluble second messengers.
Attempts have been made to group intracellular signalling proteins according to their function, but you will soon see that there are plenty that have more than one function, making classification into functional groupings difficult. Nevertheless, these descriptions give a flavour of the variety of possible signalling functions. Later in this chapter we shall discuss many examples from these groups.
Relay proteins simply pass the signal on to the next member of the chain.
Messenger proteins carry the signal from one part of the cell to another. For example, activation may cause translocation of the protein from the cytosol to the nucleus.
Amplifier proteins are capable of either activating many downstream signalling proteins or generating large numbers of second messenger molecules; they tend to be enzymes such as adenylyl cyclase, which synthesizes cAMP, or ion channels such as Ca2+ channels, which open to release Ca2+ ions from intracellular stores.
Transducer proteins change the signal into a different form. Voltage-gated Ca2+ channels are examples of signalling proteins, which fall into two of these functional categories, since in addition to their role as an amplifier protein, they detect a change in membrane potential, and transduce it into an increase in the concentration of a second messenger.
Bifurcation proteins branch the signal to different signalling pathways.
Integrator proteins receive two or more signals from different pathways, and integrate their input into a common signalling pathway.
Modulator proteins regulate the activity of a signalling protein.
Other proteins are involved purely in the correct placement of some signalling molecules:
Anchoring proteins tether members of the signalling pathway in particular subcellular locations, such as the plasma membrane or the cytoskeleton, thereby ensuring that the signal is being relayed to the right place.
Adaptor proteins link one signalling protein with the next at the correct time, without signalling themselves.
Scaffold proteins are proteins that bind several signalling proteins, and may also tether them, forming a much more efficient functional complex. Scaffold proteins may therefore share attributes of both anchoring and adaptor proteins.